1. Pre-experimental Preparations
- Prepare six solutions (see Table 1 for compositions): (a) 100 ml NZR (normal zebrafish Ringer’s), (b) 50 ml NZR + Tricaine (MS222), (c) 10 ml NZR +BSA; (d) 100 ml LoCaS (low Ca2+ solution), (e) 10 ml LoCaS + papain, (f) 100 ml K+-internal solution. Solutions (a), (b), (d) and (f) can be stored for up to one month at 4 °C. Solutions (c) and (e) should be prepared the day of experimentation. All solutions should be at room temperature before beginning experiments.
- Label and fill four 35 mm Petri dishes approximately halfway with solutions (a), (c), (d) and (e).
- Make a dissecting dish by filling a 60 mm Petri dish with Sylgard (Dow Corning, Midland, MI) and then cutting a cavity into it that will allow a fish to sit upright without tipping over.
- Prepare at least two cell isolation tools by gluing the follicle end of a dog hair to the end of a glass Pasteur pipette with superglue, allowing the hair to extend past the end of the pipette by approximately 0.5 cm. The hairs from soft-furred dogs like Labrador Retrievers and Weimaraners work well.
2. Isolation of Auditory and Vestibular Labyrinth
- Sacrifice one adult zebrafish by immersion in a beaker containing NZR + Tricaine. Visually monitor the gills until all opercular movements have ceased. Wait an additional ten minutes before removing fish and rinsing with fresh NZR. These procedures have been approved by the Institutional Animal Care and Use Committee (IACUC) of Pepperdine University but approval should be obtained from your own institution.
- Pin the fish to the dissecting dish dorsal side up by inserting one standard sewing straight pin (approximately 2.5 cm long) through each eye socket and a third pin through the dorsal-ventral axis about one-third to one-half of the distance from head to tail as illustrated in Figure 1A.
- Under a zoom (0.62-5x) stereo dissecting microscope fitted with 10x eyepieces, use spring scissors (Fine Science Tools catalogue number 15000-02) to expose the brain and a short segment of the spinal cord by removing the skull roof from a point approximately 0.5 cm caudal to the level of the gills forward to the nose of the fish. Cut the spinal cord and lift the brain with fine forceps (Fine Science Tools catalogue number 11251-30) while cutting the spinal nerves and other attachments; remove the brain and discard (Figure 1B).
- The ventral half of the skull capsule that remains forms a “bowl” that contains the inner ear organs. Observe the prominent, white, opaque otoliths in the lagenas and sacculi located symmetrically at the caudal end of each inner ear and the laterally positioned utricle otoliths more rostrally (Figures 1C and 4A).
- Remove the bowl of bone by cutting all ventral and lateral attachments while gently lifting it out of the head with fine forceps. Place it into the Petri dish containing LoCaS (Figures 1D and 4B).
- Using two pairs of fine forceps, crack apart the bowl at its midline by either grabbing the lateral edges of the bone and pulling them apart or alternatively, separating the right and left halves by inserting the points of the forceps in the center and prying the halves apart. Remove the two labyrinths from the bone using fine forceps (Figure 2).
- Inspect the labyrinths to identify the auditory and vestibular end-organs: the semicircular canals, utricles, sacculi and lagenas (Figure 4C). The pinkish areas under each otolith are the maculae containing the hair cells. Similarly, a strip of pink inside the ampullae of the semicircular canals identifies the cupula where the hair cells are located.
- Carefully remove the otoliths from the lagena and utricle using the fine forceps (Figures 2 and 4C). It is not necessary to remove the otolith from the saccule as doing so may damage the saccular hair cells.
3. Hair Cell Isolation
- Using a plastic transfer pipette (Fisher Scientific catalogue number 13-711-9AM), move the end organs to the dish containing LoCaS + papain minimizing the volume of fluid transferred.
- Incubate these structures in this solution for thirty minutes at room temperature.
- At the end of the incubation period, move the structures to the dish containing NZR + BSA and incubate in this solution for at least thirty minutes. Hair cells will remain healthy in this solution for up to four hours but will deteriorate after 1-2 hours after isolation (see next steps).
- In preparation for cell isolation, move one of the end-organs to the dish containing NZR.
- For the lagenas and utricles, use a dog hair to gently lift off the maculae-sheets of pinkish epithelial tissue that contain the hair cells (Figure 3A). The maculae can be scooped out (using a dog hair) from under the otoliths of the sacculi, and the cupulae from their locations inside the ampullae of the semicircular canals.
- Using two cell isolation tools prepared in step 1.4, triturate a macula or cupula to generate a debris field that contains the hair cells (Figure 3B). Allow the cells at least five minutes to settle onto the bottom of the dish before moving it.
- Relocate the dish to the stage of an inverted microscope equipped with phase contrast optics. Observe cells under 40X magnification and note the diversity in morphology. Many cells are avocado-shaped while others are long and thin. The presence of apical hair bundles identifies them as hair cells and phase-brightness assures their health. Photomicrographs of isolated hair cells are shown in Figure 5.
4. Patch Clamping Zebrafish Hair Cells
- In preparation for perforated patch recordings5,6, prepare amphotericin-containing internal solution as follows: place 5 mg amphotericin B (Sigma A4888) into a 1.5 ml microcentrifuge tube and fill with 100 μl DMSO. Immediately vortex this solution for 10-20 seconds until all of the amphotericin is dissolved. Then, pipette 625 μl K+-internal solution into a second microcentrifuge tube. Add 10 μl of the amphotericin solution to the K+-internal solution and vortex as above. The stock amphotericin solution will last several hours but discard the final solution after one hour.
- Fabricate patch pipettes from BF 150-86-10 borosilicate glass (Sutter Instruments) using a multi-stage puller (Sutter P-1000). Pipettes should have tip diameters of approximately 2 μm and will have resistances of 1-3 MΩ with the solutions in Table 1. The settings on the puller that work well are as follows: Heat=Ramp minus10, Pull=0, Velocity=18, Delay=1, Pressure=600. The shape of the tip of a good pipette is evident from the photograph in the inset c of Figure 5. Blunt “bullet-shaped” pipettes are preferred as they offer the least access resistance during electrophysiological recordings.
- Use a 28 gauge Microfil syringe needle (MF28G-5; World Precision Instruments) to fill a recording electrode halfway with the amphotericin-containing K+-internal solution.
- Affix the electrode to the headstage of a patch clamp amplifier. Apply -10 mV, 10 ms voltage steps at 10 Hz and maintain positive pressure in the electrode as it is maneuvered through the air/solution interface and down to the bottom of the recording dish near the cells.
- Position the electrode orthogonally to a healthy looking cell. When the electrode is near enough to the cell so that it begins move away from the out-flowing solution, quickly reverse the pressure, applying a slight vacuum to the electrode until the cell “jumps” onto it. Immediately cease suction on the electrode and apply a holding potential of -70 mV to the inside of the electrode. Within a few seconds a gigaohm seal will form.
- Observe the capacitive current transients at the onset and termination of the voltage steps that appear soon after seal formation. Continue to monitor the perforation of the membrane under the electrode by the amphotericin by watching as the transients grow in magnitude to their maximum size (in about ten minutes).
- To confirm the establishment of whole-cell (perforated) configuration in a healthy cell, elicit outward K+ currents by applying hyperpolarizing and depolarizing steps of voltage. Most hair cells have prominent outward currents (see Figure 6).
5. Representative Results
Figure 4 shows images taken during the steps of cell isolation. In panel A, the otoliths associated with the lagenas, saccluli and utricles can be seen through the thin layer of bone that overlies them. These opaque structures provide convenient landmarks to aid in the safe removal of the end organs from the animal during the subsequent steps of the dissection. Panel B shows the “bowl” of bone that contains the intact labyrinths and the prominent otoliths in the utricles, lagenas and sacculi. Panel C shows a labyrinth that has been removed from the bone. Note the large otolith with the scalloped edge in the lagena, the icicle-shaped otolith in the saccule and the bean-shaped otolith in the utricle. Some of the diversity in size and morphology seen in cells isolated from the lagena is illustrated in Figure 5. A healthy cell is readily identified as being phase bright with a sharp perimeter. Cell shapes can be roughly divided into two classes: avocado-shaped (avo) and long and thin (thn) although the sizes of cells within each group can vary markedly (compare for example avo 1 and avo 3). Inset a shows a cluster of three cells that were not completely isolated from one another. The black color and granular appearance of the cell in inset b readily identifies this cell as dead. Inset c shows the tip of a patch pipette illustrating the shape and dimensions appropriate for recording from these cells.
The current traces shown in Figure 6 were obtained in response to hyperpolarizing and depolarizing steps of potential imposed on a lagena cell similar to the avo 2 shown in Figure 5. Current responses in cells of different sizes and shapes can vary widely suggesting diversity in the complement of voltage-gated channels.
Figures 1-3: Cartoons illustrating steps in the isolation of hair cells.
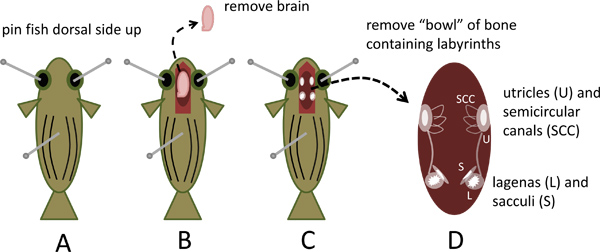
Figure 1. Removal of the skull capsule containing the inner ear labyrinths from zebrafish. A. Pin sacrificed zebrafish dorsal side up to dissecting dish. B. Open skull, remove and discard brain. C. Observe otoliths in utricles, lagenas and sacculi. D. Remove ventral portion of skull capsule containing the labyrinths.
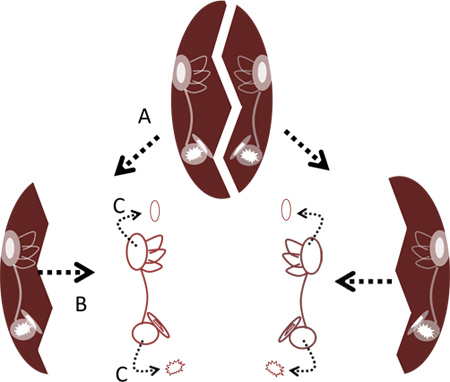
Figure 2. Removal of labyrinths from skull capsule. A. Crack apart skull capsule at its midline. B. Remove labyrinths from bone. C. Remove the otoliths from the lagenas and utricles using forceps.
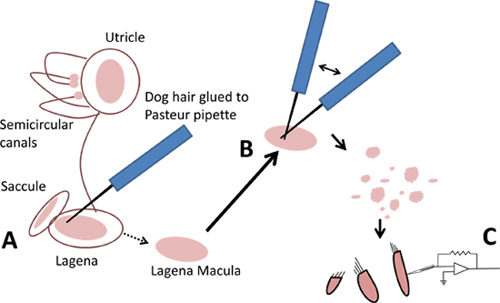
Figure 3. Isolation of hair cells from labyrinths. A. Gently lift off maculae with a dog hair. B. Triturate maculae with two dog hairs. C. Use isolated hair cells for patch clamp.
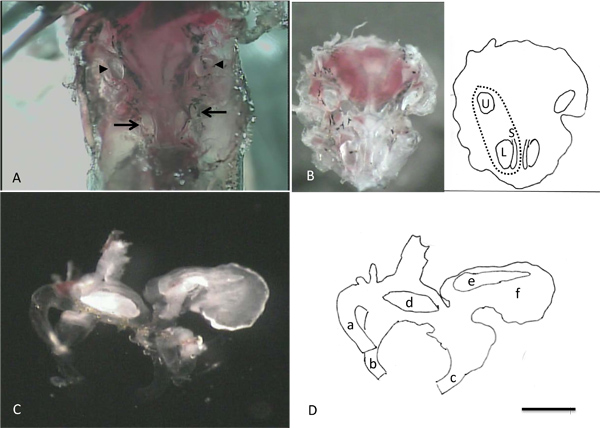
Figure 4. Images taken during steps in the isolation of individual cells. A. After removal of the brain, the otoliths associated with the two lagenas and sacculi (arrows) and utricles (arrowheads) can be visualized. B. After removal of the ventral portion of the skull capsule that contains the inner ear organs from the animal. The cartoon in the right half of B identifies the locations of the left utricle, lagena and saccule. The dashed line indicates the approximate position of the left labyrinth. C. Right labyrinth (medial view) D: Cartoon drawing illustrating key portions of labyrinth in C. a: anterior semicircular canal, b: horizontal canal, c: posterior canal, d: utricular otolith, e: saccular otolith, f: lagenal otolith. Scale bar in D represents 1 mm for A and B, 0.5 mm for C and D.
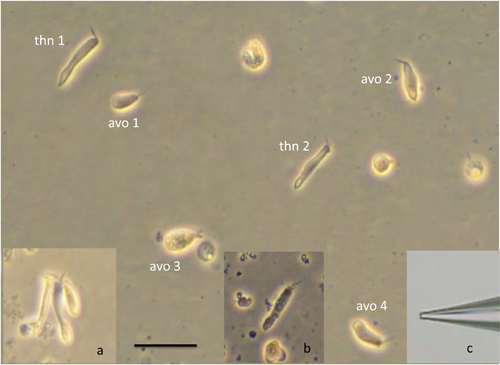
Figure 5. Isolated, healthy cells from the lagena illustrating the diversity of morphology; avo: avocado shaped cells; thn: long, thin cells. Inset a: cluster of three incompletely isolated cells; inset b: dead cell; inset c: tip of patch electrode used in electrophysiological recordings from zebrafish hair cells. Scale bar = 20 μm applies to the main figure and all insets.
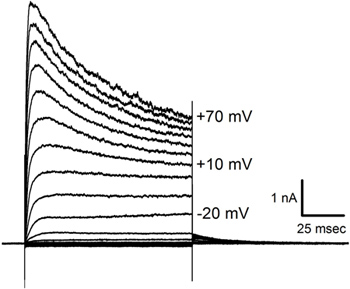
Figure 6. Currents recorded in a patch clamped hair cell. Averaged responses in a cell (similar to avo 2 in Figure 5) to three presentations of voltage steps applied in 10 mV increments from -140 mV to +70 mV from a holding potential of -70 mV. Note the inactivating current that appears at more depolarized potentials. Voltage step magnitudes are shown next to some of the traces.
