Spesialiserte spenningsklemmeteknikk tillater opptak av ioniske strømmer ved kontrollerte membranpotensialer. Mye brukt to-elektrode spenningsklemmen (TEVC) og patch clamp teknikker gir pålitelig elektrofysiologisk informasjon om egenskapene til mange ionekanaler. Men begge disse metodene har ulemper som hindrer anskaffelse av pålitelige data for rask spenningsstyrte natriumkanaler og andre fast aktiverings kanaler i membraner som for eksempel de av Xenopus oocytter. De Bezanilla og Stefani laboratorier følgelig utviklet cut-åpen vaselin gap spenning klemme teknikk (COVG) for egg 2. Teknikken har blitt brukt mye til å spille inn, Na +, K +, og Ca 2 + kanaler 3-8.
Under COVG opptak, er en heterologt protein-uttrykke eggcelle membran delt inn i tre regioner. Den ioniske strøm av data blir tatt opp fra den øverste regionen av oocytten sombad som omgir den øverste region er festet til en lede potensial, som kan enkelt og raskt skiftes. De midtre region beskytter mot lekkasjestrøm ved å bli spent fast til samme potensial som den øverste regionen ni. Den nedre område er der oocytt åpning (cut-åpen) skjer ved hjelp av et saponin løsning eller en kanyle. Kjemiske eller manuell åpning av membranen i det nedre område tillater kontroll av den indre potensial, som er festet til bakken, og gjør cellen innvendig sammenhengende med det nedre kammer løsning. Perfusjon av løsninger inn i det nedre kammer kan justere egenskapene til det indre miljø, mens oppløsningen veksling i den øverste kammeret endrer de eksterne omgivelsene. 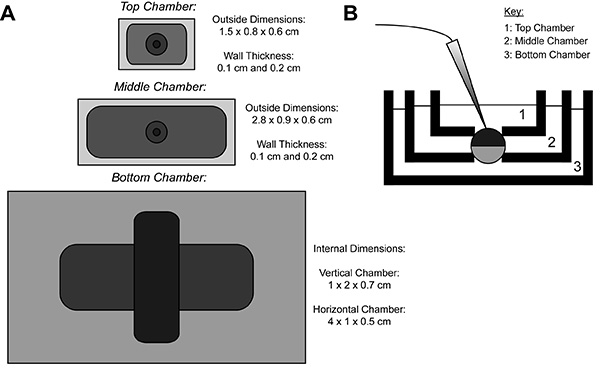
Figur 1. Eggcelle Cut-Open Voltage-Clamp Bath Setup Diagram. (A) Topned-visning av de tre bad atskilt fra hverandre. Dimensjonene av kamrene for COVG er vist på figuren. (B) Sett fra siden av badstuer oppsett i testing posisjon. Klikk her for å se større bilde .
Fordelene med COVG teknikk omfatter lav strømstøy (1 nA på 3 kHz), kontroll av den ioniske sammensetning av eksterne medier, evnen til å modulere den interne mediet, fast tidsoppløsning (20-100 usekunder tidskonstanten for nedbrytning av kapasitet transient), og stabile opptak i flere timer 9. Ulempene er at det krever spesialutstyr, og det er mer vanskelig å utføre sammenlignet med to-elektrodespenningsklem (TEVC) 10.
Mens COVG tilnærming krever høyt spesialisert utstyr og intrikate prosessuelle elementer, kan det gi rom for kjøp av verdifulledyktige elektrofysiologiske data. Denne informasjonen, som for eksempel gating strømninger med raske kinetikk og hale strømninger 4, kan tas opp uten noen av problemene forbundet med andre spenningsklem protokoller inkludert kanal nedslitt. Mindre modifikasjoner til COVG oppsett kan tillate bruk av temperaturkontrollere og spenning klemme fluorometri (VCF). Inkludering av spenning klemme fluorometri elementer innen COVG forsamlingen kan øke datautgang ved å konferere muligheten til å overvåke protein konformasjonsendringer samtidig opptak nåværende 11-13.
