This protocol allows separation of a mitochondria-containing fraction from cytosolic components. The best way to test its success is to perform northern analysis and western analysis (Figure 1) to samples from the different isolation steps. Three parameters for the isolation quality are derived from these analyses. First, whether the RNA or proteins in the samples are intact – these will be detected as distinct bands in the analyses. Degradation events will induce the appearance of additional, shorter bands or smears. Second, comparing the signals from each step with the signal from the input (i.e. Total) provides important information regarding losses during preparation. Severe losses will be observed as a significant difference between the Total and the relative amount of signal. Lastly, and most important, the quality of separation between mitochondrial and cytosolic components can be determined. Isolation of intact mitochondria will result in signal for mitochondrially-transcribed transcript only in the mitochondria fraction and not in the cytosolic fraction (Figure 1A). Furthermore, proteins that are mitochondria-associated will appear by western blot only at the mitochondria fraction, and cytosolic proteins in the cytosolic fraction (Figure 1B).
Figure 1 demonstrates two additional outcomes that are standard for this protocol: first, association of nuclear-encoded mRNAs with mitochondria varies between genes. One can see that ACO1 mRNA, which encodes a mitochondrial protein, appears mostly in the mitochondria fraction, while the mRNA encoding the cytosolic actin protein (ACT1) is mostly in the cytosolic fraction (Figure 1C). Genome-wide analyses revealed variation among many genes8,10,21,22 and the basis for this diversity is under extensive research. Second, significant amounts of ER-related material are coisolated in the mitochondrial fraction. The mRNA encoding SEC61, which is ER-associated, is detected by northern blot (Figure 1A) and the ER-membrane protein Cue1 is detected by western blot (Figure 1B). This may be an outcome of the known physical contacts between ER and mitochondria. While further purification of mitochondria is possible, it may introduce some limitations; these are discussed in the Discussion.
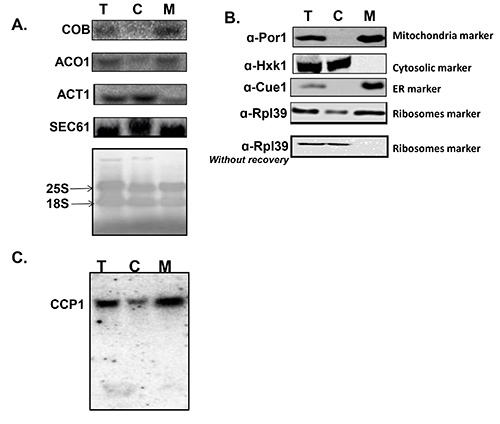
Figure 1. Quality verification of isolation. Quality is tested for samples taken in three representative steps of the procedure: Before fractionation (Total [step 1.20], T; cytosolic fraction [step 1.22], C; and mitochondrial fraction [step 1.24], M). The samples are subjected to northern analysis (A, C) or western analysis (B). A) Northern analysis for the following mRNAs: COB is an RNA that is transcribed inside the mitochondria therefore its signal should be exclusively in the mitochondria. Its appearance in the C fraction will indicate of mitochondria lysis during preparation. ACT1 is an mRNA that encodes a cytosolic protein, and therefore translated by cytosolic ribosomes and appears mostly at the C fraction. ACO1 is an mRNA that encodes a protein that is imported to the mitochondria, and appears mostly at the mitochondria fraction. SEC61 is an mRNA that encodes ER resident protein; its presence in the M fraction indicates the presence of ER components in this fraction. The bottom panel presents a methylene blue staining of the northern membrane, is which two rRNAs (18S and 25S) are detected. This panel demonstrates that significant amount of ribosomes appear in the M fraction. B) Western analysis for the following proteins: Por1 is a mitochondria outer membrane protein therefore its signal is expected only in the M fraction. Signal in the C fraction will suggest dissociation of mitochondrial components during preparation. Hxk1 is a cytosolic protein and Cue1 is an ER protein. Both report on the copurification of these fractions with the mitochondria fraction. Rpl39 is a ribosomal protein, again demonstrating the presence of ribosomes in both fractions. The panel “Rpl39 without recovery”, presents the fractionation of ribosomes when the protocol is excluded of the recovery step (step 1.14). C) Northern analysis for CCP1, an mRNA that encodes a protein that is imported to the mitochondria. The entire gel is presented to demonstrate the lack of shorter, degradation-originated bands. Please click here to view a larger version of this figure.
