The maturation stage of parasites prior to E64 treatment is critical for generating membrane-enclosed merozoites. Figure 1Ai shows the appropriate maturation stage of schizonts for adding E64. The parasites should be large and almost fill the red blood cell. A dappled appearance of Giemsa stain indicates merogony has commenced, and E64 should be added to yield membrane-enclosed merozoites (Figure 1Aii). If E64 is added earlier to trophozoite stage parasites, membrane enclosed merozoites are not generated even after 12 hr of E64. Instead parasites take on abnormal morphology such as enlargement of the digestive vacuole (Figure 1Bi and 1Bii), and merozoites are not formed. If E64 is added later to schizonts, membrane-enclosed merozoites are not generated as schizont rupture is uninhibited (Figure 1Ci and 1Cii). A high level of parasite synchrony is required, otherwise a range of all three outcomes described will be seen after E64 treatment.
Removal of hemozoin is another critical step for purifying merozoites. If merozoites are not purified from hemozoin then merozoite-hemozoin clusters form which cannot be dislodged by pipetting. These aggregates run on the cytometer as single events, albeit with different forward-scatter and side-scatter characteristics than single merozoites, resulting in inaccurate merozoite counting by flow cytometry (Figure 2A). As THP-1 cells can also phagocytose hemozoin, these merozoite-hemozoin clusters can also be phagocytosed (Figure 2B). These aggregates are highly EtBr fluorescent as they contain multiple merozoites. Phagocytosis of aggregates results in a THP-1 EtBr fluorescence profile equivalent to that observed for the phagocytosis of multiple individual merozoites. Hence, if hemozoin is not removed, the EtBr fluorescence of THP-1 cells will be an overestimate of the opsonizing potential for the plasma being tested. Hemozoin is also reported to significantly alter phagocyte function. GFP positive merozoites are counter stained with EtBr to increase visualization of merozoites phagocytosed by THP-1 cells. Using the conditions described in this protocol, all GFP positive merozoites are counterstained with EtBr which has a brighter fluorescence intensity (Figure 3A). Phagocytosis assessment by EtBr fluorescence enables superior resolution of merozoite phagocytosis than GFP fluorescence (Figure 3B).
The number of merozoites added to the assay will modulate the amount of phagocytosis observed. For this reason, accurate counting by flow cytometry is critical. Increasing ratios of merozoites:THP-1 will result in increased adherence of merozoites to THP-1 cells in the absence of plasma (Figure 4A). Increasing merozoite:THP-1 ratios also results in increased phagocytosis responses when merozoites are opsonized (Figure 4B). A 4:1 merozoite:THP-1 ratio is recommended for robust phagocytic responses. Using this ratio, and the dilution of plasma indicated, this assay is able to resolve low, intermediate and high levels of opsonizing antibodies. Between 0 and 78 percent phagocytosis responses have been observed using PNG plasma samples and the other conditions described. Such responses are recently shown to associate with naturally acquired clinical immunity to malaria10. Figure 5 shows examples of the 4 quartiles of phagocytosis responses from PNG individuals.
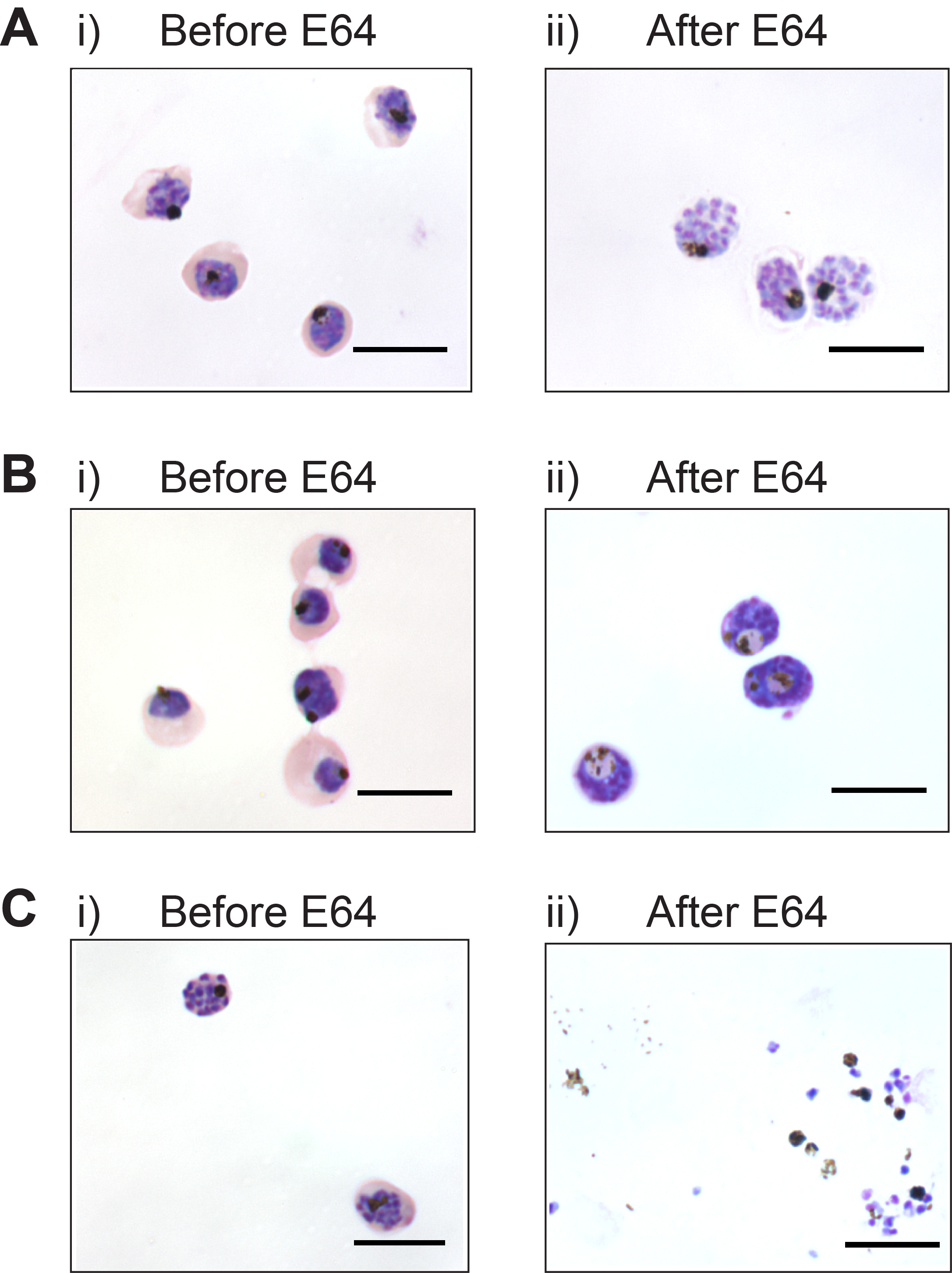
Figure 1. Timing of E64 addition is critical for generating membrane-enclosed merozoites. (A) Appropriate maturation stage of parasites i) prior to E64 treatment, and ii) membrane-enclosed merozoites produced after 6 hr of E64 treatment. (B) membrane enclosed merozoites are not generated if E64 is added to immature parasites; i) Late stage trophozoites, and ii) after 12 hr of E64. (C) Schizont rupture is not inhibited if E64 is added to late segmented schizonts; i) Late stage schizonts, and ii) after 6 hr of E64. Scale bar represents 10 μm.
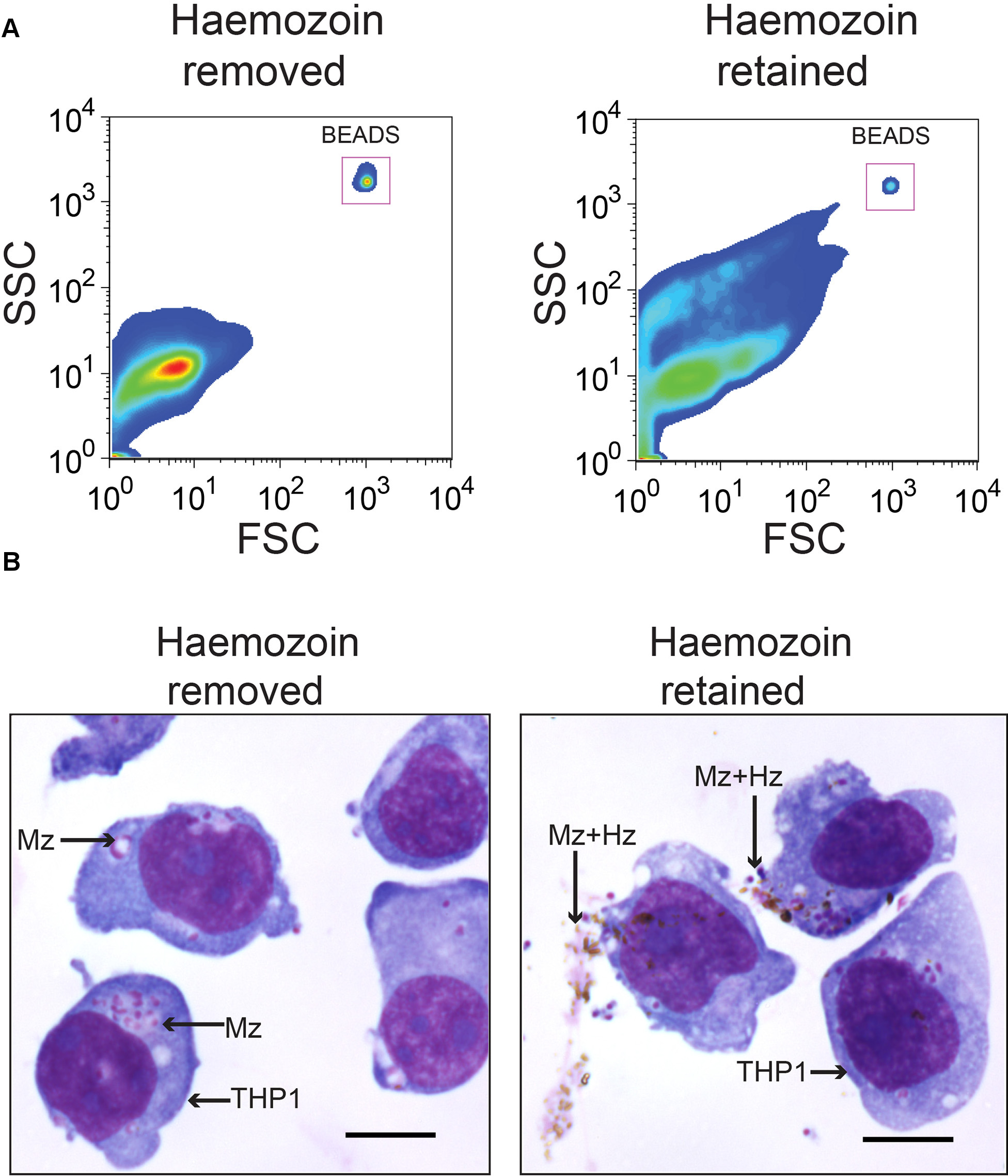
Figure 2. Hemozoin removal is critical to generate a single-cell merozoite suspension. Merozoite-hemozoin aggregates form following filtration of membrane enclosed merozoites, unless hemozoin is magnetically separated from merozoites. (A) Forward-scatter and side-scatter plots of merozoites with hemozoin removed and hemozoin retained. (B) Hemozoin-merozoite aggregates can be phagocytosed by THP-1 cells. Diff-Quick stained cyto-spin slides of THP-1 cells incubated with PNG plasma opsonized merozoite preparations with or without hemozoin. Scale bar represents 10 μm.
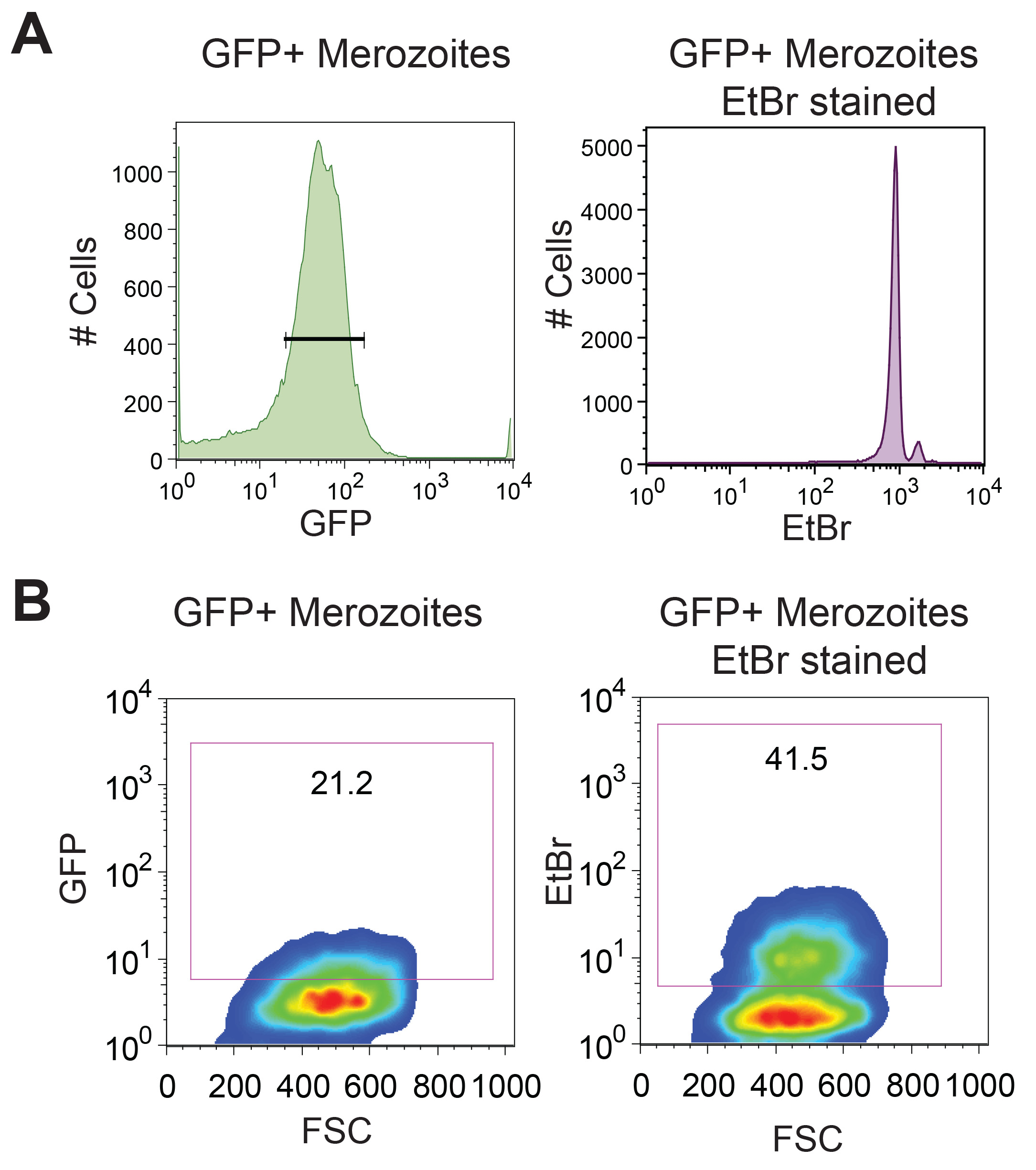
Figure 3. Ethidium Bromide staining allows for superior resolution of merozoite phagocytosis. D10-GFP purified merozoites are counterstained with EtBr to improve fluorescence intensity. (A) Flow cytometry histrogram of purified merozoites with gating on GFP positive merozoites, and a dot blot showing EtBr fluorescence within this gated GFP positive population. (B) Forward scatter versus GFP and forward scatter versus EtBr of THP-1 cells following phagocytosis of GFP and EtBr dual fluorescent merozoites. Gates were drawn based on the THP-1 cells with merozoites and no plasma control.

Figure 4. The merozoite:THP-1 ratio influences the degree of phagocytosis observed. (A) Five merozoite:THP-1 ratios are depicted; 50:1, 20:1, 10:1, 4:1, 1:1. Cells only control indicates background fluorescence due to THP-1 cells. THP-1 EtBr fluorescence for each ratio is indicated for merozoites opsonized with a pool of PNG plasma or with nonimmune Australian plasma. (B) Mean fluorescence intensity of THP-1 cells for different merozoite:THP-1 ratios, and opsonization with a pool of PNG plasma or nonimmune Australian plasma (mean+SEM). Please click here to view a larger version of this figure.

Figure 5. A large range of opsonizing antibodies can be measured. Merozoites were opsonized with plasma from PNG individuals and incubated with THP-1 cells. Four representative phagocytosis responses are shown. Gates were drawn based on the THP-1 cells with merozoites and no plasma control, and numbers indicate the %phagocytosis after subtraction of the THP-1 cells with non-immune plasma control. Please click here to view a larger version of this figure.
