The key premise of freeze-fracture image interpretation is that fracture planes pass through the lipid bilayer of membranes conferring two fracture faces, called by convention the PF-face (plasma fracture-face) and EF-face (extracellular fracture-face) (Figure 3). The PF-face is the half of the membrane lipid bilayer adjacent to the cytoplasm of the cell and the EF-face is the half of the membrane lipid bilayer adjacent to the extracellular milieu. The freeze-fracture technique is particularly useful for the investigation of membrane structure and the two faces are typically distinctively different with PF-faces being populated with many membrane associated particles in contrast to EF-faces which contain fewer. The cytoplasmic contents of freeze-fractured cells are typically coarse in appearance and are not particularly revealing although membrane bound organelles such as nuclei and Golgi can be readily identified. Of particular interest to investigators using this technique are specializations of membrane structure that can be interpreted for their function. These include specific distributions of membrane associated particles, ciliary membrane specializations, and intercellular junctions.
Cilia-associated Structures
Residing in the cell membrane at the base of each eukaryotic cilium and flagellum is an array of membrane associated particles organized into strands encircling the shaft of the cilium and known as the ciliary necklace (Figure 8)12-16. This structure exhibits some degree of morphologic heterogeneity in various species and it has been speculated to be multifunctional inasmuch as it appears prior to the emergence of the ciliary shaft during ciliogenesis and remains in place in the mature cilium. The nascent ciliary necklace derives from the aggregation and organization of membrane particle arrays prior to the time of the emergence of the ciliary shaft during ciliogenesis suggesting its possible participation in early organization of the developing axoneme14.
Tight Junctions and Epithelial Permeability
In various epithelia, freeze-fracture studies reveal a sophisticated organization of anastomosing strand and groove structures encircling the apical aspect of the basolateral borders. This belt-like structure is known as the tight junction or zonulaoccludens (Figure 9) and is believed to act as a regulator of epithelial permeability to the paracellular flux3, 17-20. Tight junctions with few strands have been described as “leaky” while those with more complex organization are described as “tight”. This organization appears to fit the function of the epithelia to which they are associated. The PF-faces of tight junctions appear as strands while the EF-faces exhibit complementary grooves.
Gap Junctions and Intercellular Communication
Gap junctions (Figure 10) appear in freeze-fracture preparations of a variety of tissues and are represented by dense arrays of particles on PF-faces and complementary pits on EF-faces. These structures are thought to represent membrane sites that facilitate intercellular communication. In tissues where they are present, demonstrations of the passage of low molecular weight fluorescent dyes and calcium fluxes have been demonstrated21-24 suggesting that they represent a physical route for the passage of signaling molecules.

Figure 1. Layout of a commercial freeze-fracture/etch plant. At far left is a pressurized Dewar feeding cooling liquid nitrogen to the specimen stage and cooling microtome arm under controlled conditions. At center is the unit with specimen chamber and electronic control panels to the right. At right is an additional liquid nitrogen tank on which the vent gas is used to bring the specimen chamber to atmosphere.

Figure 2. Freeze-fracture/etch plant specimen chamber with door opened to reveal positions of directionally aimed electron guns, cooling shroud, and specimen post where specimens in brass holders are positioned. Specimens are introduced to the chamber through a port to the left of the chamber.
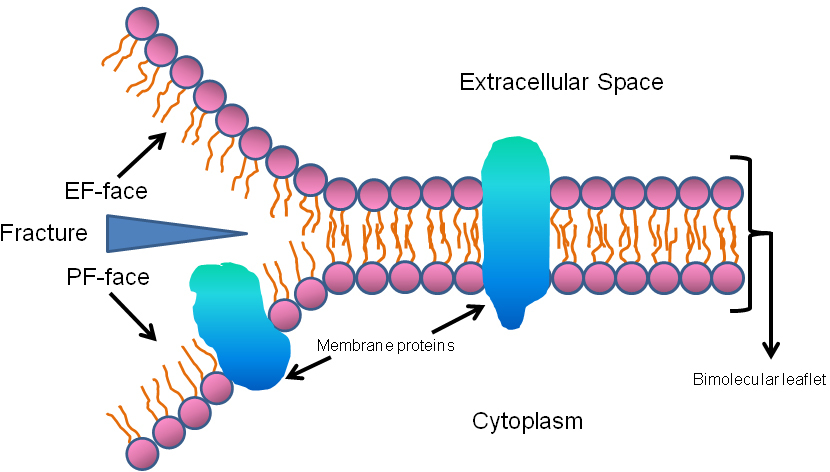
Figure 3. Diagrammatic representation of the principle of freeze-fracture. The fracture faces derive from the splitting of the membrane through the lipid bilayer producing an extracellular face (EF-face) and a face that is proximal to the cytoplasmic aspect, the PF-face. True unfractured membrane surfaces are seen only by etching subsequent to the freeze-fracture maneuver.

Figure 4. Two types of specimen mounts for freeze-fracture/etch procedures. (Left) A double replica specimen holder booklet is used for conventional freeze-fracture procedures performed without etching. The specimen is sandwiched between two gold specimen stubs and positioned in the brass booklet. The specimen is fractured by opening the booklet under vacuum in the specimen chamber (See freeze-fracture_movie1.mov). (Right) A mount used primarily for freeze-etching. The specimen is positioned on the gold stubs and locked into the brass holder. The specimen is fractured by gentle shaving with a razor blade fixed in the microtome arm. This configuration is preferred for etching procedures (See freeze-etch_movie2.mov).

Figure 5. Preparation of a freeze-fracture specimen mount. The specimen is sandwiched between two gold mounts (A) and frozen in a well containing liquid nitrogen cooled propane (foreground) with subsequent transfer to a holding Dewar containing liquid nitrogen (background) (B).

Figure 6. Demonstration of positioning of a frozen freeze-fracture specimen into the double replica specimen holder booklet in preparation for introduction to the specimen chamber of the freeze-fracture/etch plant.

Figure 7. Retrieval of replicas following fracture and shadowing in the freeze-fracture/etch plant. “A” is a clean water surface in a spot plate. The replica floated from the specimen stub is transferred to “B” a well containing an acidified solution of sodium dichromate or full strength household bleach for digestion of the tissue from the replica. “C” represents subsequent clean water surfaces to which the cleaned replica is transferred prior to being retrieved onto a copper electron microscopy grid.

Figure 8. Cilia related structures revealed by freeze-fracture. White arrows point to specialized membrane particle arrays on the luminal border of epithelial cells undergoing ciliogenesis. These particle arrays represent nascent ciliary necklaces which reside at the bases of emergent and mature cilia (black arrow). Please click here to view a larger version of this figure.

Figure 9. Fracture through epithelial cell membranes revealing examples of PF- and EF-fracture faces. The strand and groove organization of tight junctional complexes is evident and the intercellular space is marked by arrows. Please click here to view a larger version of this figure.

Figure 10. A freeze-fracture image of a gap junction in rat liver. Particles are evident on the PF-face and complementary pits are evident on the EF-face. Please click here to view a larger version of this figure.

Figure 11. Immunohistochemical localization of a connexin using a colloidal gold labeled antibody on a freeze-fracture preparation illustrating specific localization on the gap junction. Please click here to view a larger version of this figure.

Figure 12. An example of rapid freezing with deep etching and rotary shadowing. The epithelial cell has been fractured through the cytoplasm and a PF-face is evident (arrowheads). Behind the arrowheads can be seen a true cell surface revealed by subsequent etching. Please click here to view a larger version of this figure.

Figure 13. An example of rapid freezing with deep etching and rotary shadowing to reveal molecular complexes comprising a bovine tracheal ciliaryaxoneme. Please click here to view a larger version of this figure.
