1. Hydra Culture and Measurement of the Feeding Response
- Maintain hydra polyps in culture by feeding them daily with artemia and keeping them in a medium (1 mM Tris-HCl buffer, pH 7.6, 1 mM NaCl, 1 mM CaCl2, 0.1 mM KCl, and 0.1 mM MgSO4) contained in a glass bowl at 18 °C under 12 hr light-12 hr dark cycles as described earlier12.
- For measuring the feeding response, transfer one mature hydra polyp having 5 to 6 tentacles to a single well of a 24-well plate. Remove the residual medium from the well by tilting it, and then immediately add 500 µl of fresh medium.
- Prepare 9 µM glutathione solution in hydra medium. Since the glutathione solution is prone to oxidation, always use freshly prepared glutathione solution for each experiment.
- Transfer the plate to the imaging platform of a microscope having provisions for image recording. Use a dark background such that the behavior of hydra polyp can be clearly imaged against the contrasting background.
- The room used for observing and imaging the behavior of hydra free from lights of fluctuating intensities, air currents and noise. Such disturbances could also cause the hydra polyp to show contraction of the tentacles – even in the absence of glutathione.
- Allow the polyp to relax for 5 min.
- Make sure that the polyp is located along the central region of the well such that the behavior can be imaged clearly. If the polyp is at the edge of the well, bring it to the center by flushing the medium using a pipette and again allow it to relax.
- Capture an image of hydra in the relaxed state. This will be the zero-time point observation.
- Quickly add 9 µM glutathione solution to reach a final concentration of 3 µM in the well. Depending on the purpose of the experiment and the response shown by hydra, test a range of different concentrations of glutathione and choose the appropriate concentration required.
- Start the timer immediately after adding glutathione solution and capture images after every 15-30 sec for 4-5 min. Do not change the magnification settings during the time-lapse imaging.
- Add the glutathione solution gently and with uniform flow such that the position of the animal in the well would be minimally disturbed in the field of view of the microscope. However, if the polyp moves extensively after adding glutathione solution, move the plate very gently to bring the polyp in the field of view for image capture.
- In the control experiment, use medium lacking glutathione while keeping all the other parameters identical.
- Make sure to perform all of the above experimental steps during the first half of the day – before 1 P.M. to avoid the possible effect of circadian rhythm on the extent of feeding response.
- Open each of the captured images using the GNU Image Manipulation Program (GIMP).
- Use the “Measure” tool available from Menu > Tools > Measure to determine the distance between apical end of each of the tentacles and hypostome. If the mouth opening is observed in any of the images, determine the distance between the center of the opened mouth and the apical end of the tentacle. Refer to this distance as the tentacle spread.
- Calculate the average tentacle spread for each polyp before and after glutathione exposure. Calculate the ratio of average tentacle spread at zero-time point to that at each of the subsequent time-points. This ratio will be called relative tentacle spread.
- Repeat measurements for at least 20 polyps.

2. Method Validation using the Starvation Model
- For starvation, transfer a few hydra polyps to a separate glass bowl and do not feed them for 5 days. Feed the control group of a few polyps daily with artemia in a similar-sized bowl. Change the medium from both experimental bowls daily to avoid fungal growth in the medium.
- On the day of the experiment, feed the control group of hydra with artemia for 1 hr and use these hydra for the subsequent experiments after removing all uneaten and dead artemia from the medium.
- Measure the feeding response of the starved hydra in comparison with the hydra from control group by the method described earlier in step 1. To avoid any bias due to time of observation, alternate the measurements of each of the starved and control hydra polyps.
Glutathione causes hydra to exhibit curling of the tentacles towards the mouth for the purpose of engulfing the prey. Such curling of tentacles brings apical ends of the tentacles closer to the hypostome. This results in the reduction in the tentacle spread, or the linear distance between apical end of the tentacle and hypostome (Figure 1). The relative tentacle spread, or the ratio of average tentacle spread before and after adding glutathione, averaged across multiple polyps reduces over time. The relative tentacle spread after addition of the medium lacking glutathione, however, reduces only transiently and attains unit value in about a minute (Figure 2). Starvation is known to cause enhancement of the feeding behavior induced by glutathione1,13. The method of measuring the feeding behavior described here was validated by the starvation model. The tentacle spread one minute after adding glutathione was significantly lower for the starved group as compared to the satiated group (p = 1 x 10-16 by applying two tailed t-test with equal variance) (Figure 3).

Figure 1: Method of measuring average tentacle spread. An image of a relaxed hydra polyp was recorded before adding GSH and the linear distance between mouth and apical end of each of the tentacle of the polyp was measured in terms of number of pixels using the GIMP tool. Another image was captured after adding GSH and the tentacle spread was measured in similar manner.
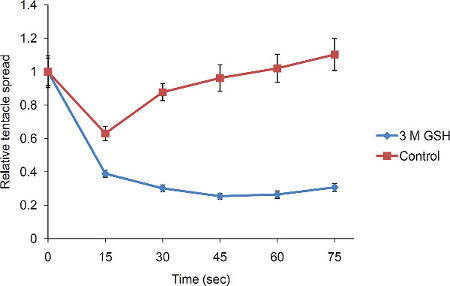
Figure 2: Quantitation of the GSH-induced feeding response in hydra. Tentacle spread at different time-points after adding GSH was measured as described in Figure 1 and relative tentacle spread was denoted as a ratio of tentacle spread before and after adding GSH (n = 14). The values for the control group represent the relative tentacle spread after addition of hydra medium without GSH (n = 14). Errors bars represent ± SEM.
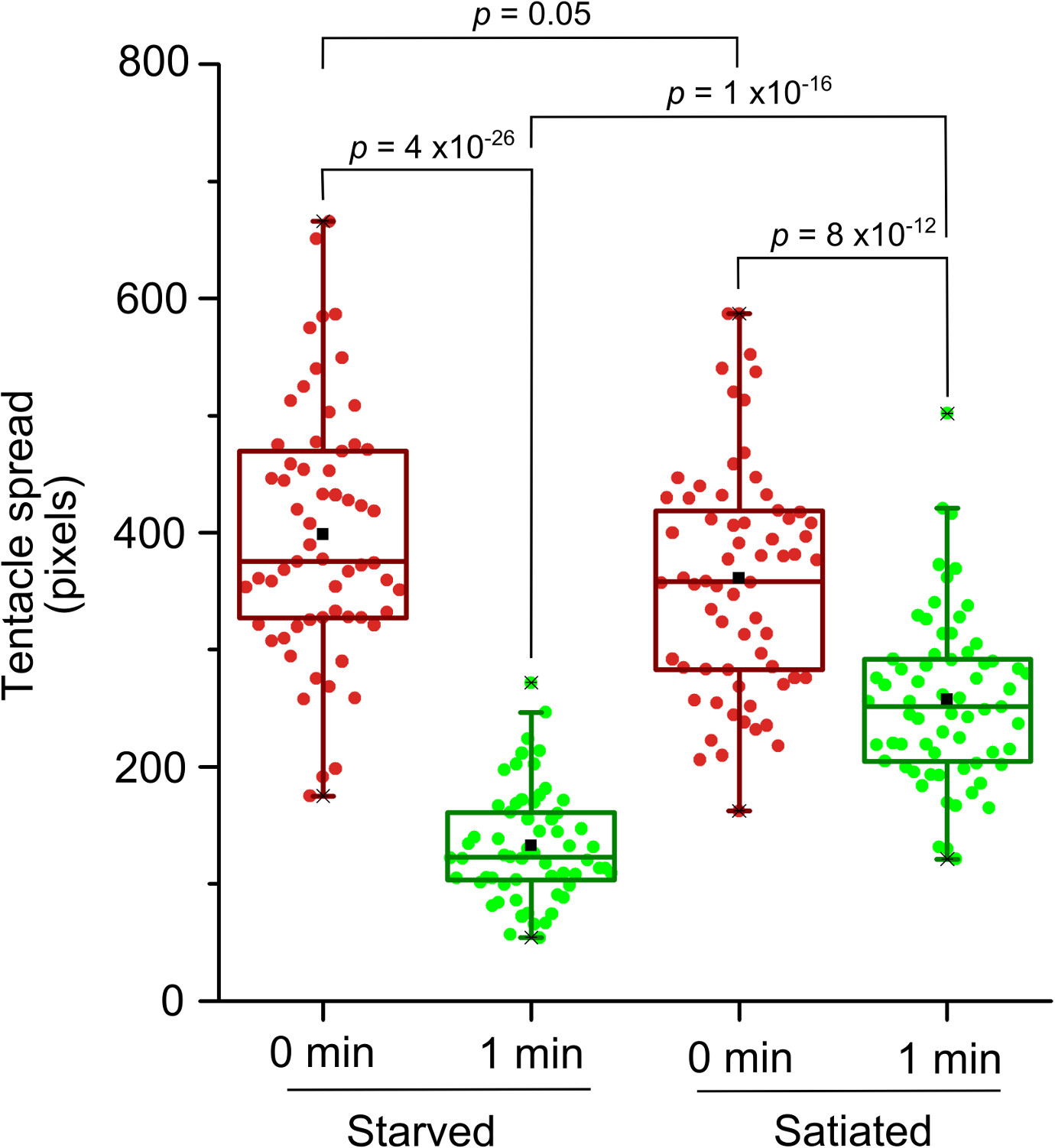
Figure 3: Validation of the method of quantitation of the feeding response using starvation model. X-axis represents the time after addition of 3 µM GSH for starved (n = 61) and satiated (n = 66) groups.
