Initially, we analyzed the effect of stimulating EC with TNFα on the recruitment of neutrophils from flow using the Ibidi microslide model (Section 7 – 9). In the absence of TNFα, little if any neutrophils adhered to the endothelial monolayer (Figure 2A). This was expected, as untreated/resting EC do not express the necessary adhesion molecules (selectins) or chemokines to support binding25,26. In contrast, cytokine-stimulation significantly increased neutrophil adhesion to the endothelium in a dose-dependent manner (Figure 2A). Adhesion normally remains stable over the course of the assay. Binding of leukocytes to untreated endothelium indicates that either the EC are activated (i.e., contaminated with LPS during the culture process) and/or the neutrophils were activated during the isolation process. Indeed, LPS has been shown to increase the expression of E-selectin, ICAM-1 and VCAM-125-27 on the surface of EC, allowing them to bind neutrophils.
When analyzing the behavior of the recruited neutrophils we typically observe a dose-dependent decrease in the percentage of neutrophils rolling (Figure 2B) with a concomitant dose dependent increase in the percentage of neutrophils migrating through the endothelial monolayer (Figure 2C). At the lower doses of TNFα-stimulation (1 U/ml) a larger proportion of neutrophils appear phase bright indicating that they are attached to the apical surface of the endothelium (Figure 2B). In contrast, at 10 and 100 U/ml (higher doses) approximately 40% of the recruited neutrophils appear phase dark at 2 min indicating that these cells have migrated through the endothelial monolayer and are underneath the endothelium (Figure 2C). Neutrophils are able to migrate through the EC within 1 – 2 min, with transmigration reaching maximal levels at ~10min post-perfusion28. Here we observed an increase in neutrophil transmigration from 40% at 2 min to 60% by 9 min post perfusion (Figure 2C). We observed no effect of TNFα concentration on the velocity of rolling (~3 µm/sec) or migrating (~10 – 12 µm/min) neutrophils.
In the filter based model, TNFα-stimulation increased neutrophil adhesion in a dose-dependent manner similar to that seen using the Ibidi microslide model (Figure 3A). In terms of behavior, neutrophil rolling was unaffected by TNFα dose (Figure 3B), whilst a dose-dependent increase in percentage transmigration was observed (Figure 3C). In this series of experiments we observed no significant effect of time on neutrophil transmigration (Figure 3C).
We provide methods on how to generate two different co-culture constructs, each of which is devised to answer specific questions. In the Ibidi microslide model, EC and MSC are cultured in a single monolayer in direct contact with one another. This model is useful for examining the effect of therapeutic injection of MSC into the blood and their subsequent integration into the EC monolayer. In contrast in the filter-based model, EC and MSC are cultured on opposite sides of the filter in close proximity but not necessarily in direct contact. This more closely resembles tissue, with endothelial cells forming a monolayer representing the blood vessel, and MSC residing in the subendothelial compartment. This allows us to examine the effects of tissue-resident MSC on the response of EC to cytokine stimulation.
Based on our experiences, we observe that maximal neutrophil recruitment and transendothelial migration occurs when EC are stimulated with 100 U/ml TNFα. As such, we have used this concentration to examine the effect of MSC co-culture on endothelial recruitment of neutrophils from flow. Here, we present data for BMMSC in co-culture with EC using the microslide and filter-based models however, other types of MSC can also be examined e.g., WJMSC. In both models the presence of BMMSC significantly reduced neutrophil adhesion to the EC when compared to EC cultured alone (Figure 4A). Co-culture had no effect on the behavior of recruited neutrophils, with similar levels of rolling and transmigration observed on EC cultured alone or with MSC (Figure 4B and C). Thus, MSC can modify the EC response to cytokine stimulation, which suppresses their ability to support neutrophil recruitment from flow.
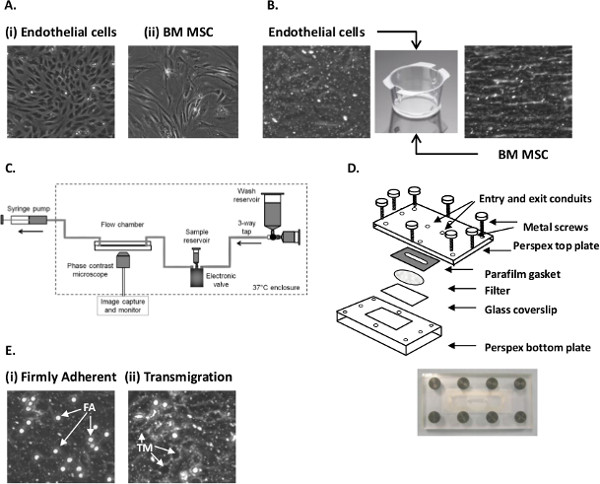
Figure 1. Establishing EC-MSC co-culture and analyzing neutrophil recruitment using a flow-based adhesion assay. (A) Micrograph of (i) primary EC and (ii) passage 3 BMMSC grown on tissue culture flasks. (B) Micrograph of EC and MSC cultured on 6-well Transwell filter inserts. (C) Diagram of the perfusion system used to generate flow. (D) Schematic representation of the parallel plate filter flow chamber. (E) Micrograph of (i) firmly adherent (FA) and (ii) transmigrated (TM) neutrophils following recruitment from flow to EC stimulated with 100 U/ml TNFα. Images C and D are taken from Figures 2 and 3 in Methods in Molecular Biology: T-cell Trafficking, 2010, pg 53-5428 with kind permission from Springer Science and Business Media. Please click here to view a larger version of this figure.
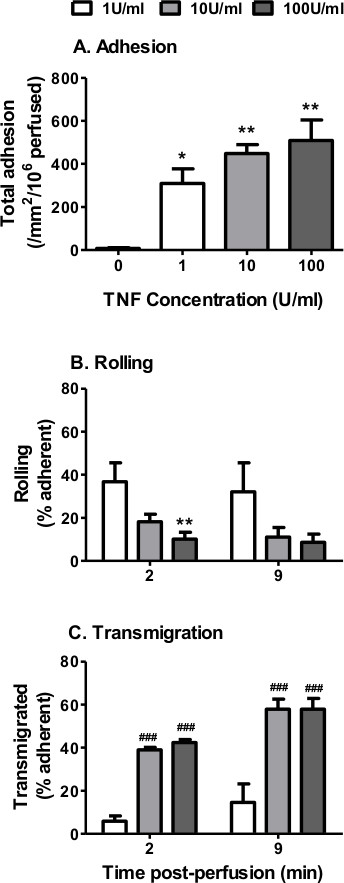
Figure 2. Neutrophil recruitment from flow to TNFα-stimulated EC using Ibidi microslides. EC were stimulated with increasing concentrations of TNFα (0 – 100 U/ml) for 4 hr. A 4 min bolus of neutrophils was perfused over the EC monolayer at 0.05 Pa. (A) Neutrophil adhesion assessed at 2 min. ANOVA showed a significant effect of TNFα treatment on neutrophil adhesion, p < 0.01. Neutrophil behavior was assessed at 2 and 9 min and expressed as a percentage of adherent cells that were (B) rolling or (C) transmigrated. ANOVA showed a significant effect of TNFα treatment on the behavior of the adherent neutrophils, p < 0.001. In C, ANOVA showed a significant of time on transmigrated neutrophils p < 0.01. Data are mean ± SEM from n = 3 experiments. *p < 0.05 and **p < 0.01 compared to the unstimulated (0 U/ml) EC control by Dunnett post-test. ##p < 0.01 and ###p < 0.001 compared to the 1 U/ml EC at the same time point by Bonferroni post-test.
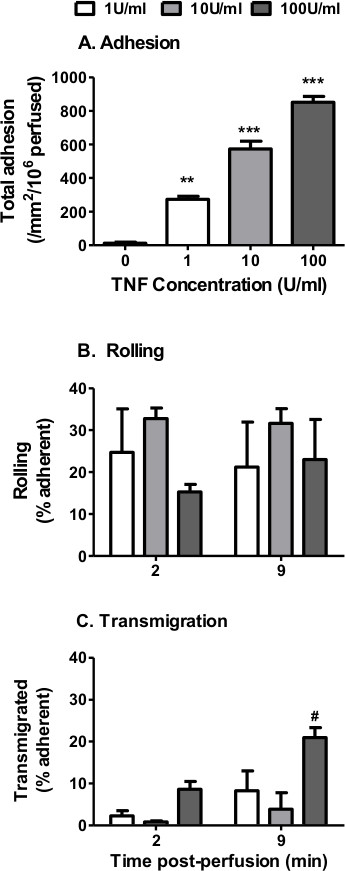
Figure 3. Neutrophil recruitment from flow to TNFα-stimulated EC using filter-based assay. EC were stimulated with increasing concentrations of TNFα (0 – 100 U/ml) for 4 hr. A 4 min bolus of neutrophils was perfused over the EC monolayer at 0.1 Pa. (A) Neutrophil adhesion assessed at 2 min. ANOVA showed a significant effect of TNFα treatment on neutrophil adhesion, p < 0.001. Neutrophil behavior was assessed at 2 and 9 min and expressed as a percentage of adherent cells that were (B) rolling or (C) transmigrated. In C, ANOVA showed a significant effect of time and cytokine treatment on neutrophil transmigration, p < 0.05. Data are mean ± SEM from n = 3 experiments. **p < 0.01 and ***p < 0.001 compared to the unstimulated (0 U/ml) EC control by Dunnett post-test. #p < 0.05 compared to the 1 U/ml EC at the same time point by Bonferroni post-test.
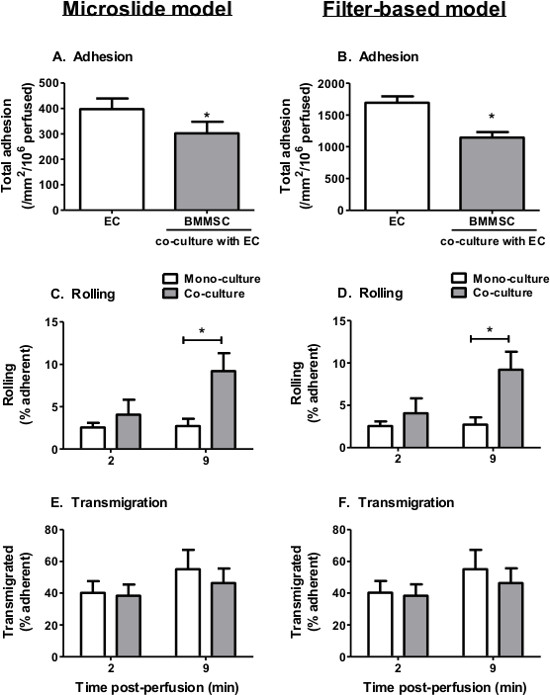
Figure 4. Neutrophil recruitment from flow to TNFα-stimulated EC-BMMSC co-cultures. BMMSC were co-cultured with EC for 24 hr prior to stimulation with 100 U/ml TNFα for 4 hr. A 4 min bolus of neutrophils was perfused over the EC monolayer at 0.05 Pa for (A, C, E) microslides and 0.1 Pa for (B, D, F) filters. (A-B) Neutrophil adhesion was assessed at 2 min. Neutrophil behavior was assessed at 2 and 9 min and expressed as a percentage of adherent cells that were (C-D) rolling or (E-F) transmigrated. In C and D, ANOVA showed a significant effect of culture conditions on neutrophil rolling, p < 0.05. In E and F, ANOVA showed a significant effect of time on neutrophil transmigration, p < 0.05. However, no significant differences were observed in transmigration between individual treatments by Bonferroni post-test. Data are mean ± SEM from n = 5 experiments. *p < 0.05 compared to the EC monoculture by paired t-test or Bonferroni post-test.
Supplementary Video 1. Analysis of neutrophil rolling velocities. EC cultured on a Transwell filter were stimulated with 100 U/ml TNFα for 4 hr. A bolus of neutrophils was perfused over the EC for 4min. Representative digitized sequence of a single 10 sec field taken 2 min post-perfusion. The change in position of a single rolling neutrophil from the beginning to the end of the 10 sec sequence can be used to calculate the velocity at which the neutrophil is rolling.
Supplementary Video 2. Analysis of neutrophil migration velocities. EC cultured on a Transwell filter were stimulated with 100 U/ml TNFα for 4 hr. A bolus of neutrophils was perfused over the EC for 4 min. Representative digitized sequence of a single 5 min field to track the movement of transmigrated neutrophils. This can be used to calculate the velocity.
