In Figure 1, the schematic demonstrates the negative panning process by which we removed phage binding off-target antigens from our library using immunotubes. We initially started with BSA since this is a common blocking agent and any phage that would react nonspecifically with this target would be problematic in future immunoassays. Next, we removed binders to aggregated alpha-synuclein to eliminate phage that are reactive with generic structures of aggregated proteins (i.e., an antibody that would be cross-reactive to aggregated alpha-synuclein, TDP-43, abeta, etc.). AFM results showed phage binding after 1 round of negative panning against the aggregated alpha-synuclein (Figure 2A) and no binding after 8 rounds (Figure 2B). We then negatively panned against healthy human brain tissue to remove phage binding the many antigens present in healthy human brain homogenate. Figure 2C showed phage binding to healthy tissue after 1 round of negative panning and no phage left binding after 10 rounds of negative panning (Figure 2D). Since the amount of healthy and FTD immunoprecipitated TDP-43 protein available for panning was low, using mica rather than immunotubes utilized less volume and therefore lowered total protein consumed (Figure 3). After 8 rounds of negative panning against healthy immunoprecipitated TDP-43, the phage was divided. Half the phage was expended in two rounds of negative panning against FTD TDP-43. The objective was to eliminate any FTD TDP-43 reactive clones (due to TDP-43’s involvement in FTD), while retaining any potential ALS TDP-43 specific clones.
For the positive panning portion (Figure 4), we also employed mica as a substrate to minimize use of material. We used the unbound phage after the FTD TDP-43 negative panning against ALS TDP-43 to acquire any ALS specific clones. We also positively panned against ALS TDP-43 twice in the event that we did not obtain clones after using the unbound phage from the negative panning against FTD TDP-43. After the entire panning process, our three elution methods yielded 154 clones from the two positive panning against ALS TDP-43 (clones that may be reactive to ALS and FTD) and 45 potential ALS specific clones (using the unbound phage from the FTD TDP-43 negative panning).
To further reduce the 45 potential ALS specific clones, the clones are sequenced and only the clones without any stop codons were considered further, except for one clone which showed up twice. This left us with 23 potential ALS specific clones. After preparing both phage and soluble scFv with these clones, they are tested in indirect ELISAs for specificity to ALS tissue. Almost all of the phages showed a preference for ALS tissue over healthy tissue (Figure 5). Similar results are obtained when comparing phage binding to ALS to FTD tissue (Figure 6). For future studies it is essential that these clones express high yields of functional scFvs, so we produced small batches of the different scFvs and carried out similar indirect ELISAs. Comparison of scFv binding to ALS tissue to both healthy (Figure 7) and FTD tissue (Figure 8) again showed selective binding to ALS tissue in almost all clones.
We used another immunoassay (dot blots) to further verify binding to ALS tissue and also to ascertain which clones are reactive in other immunological applications. Clone 2A binding to ALS tissue is shown (Figure 9). All of these clones showed promising results in either or both of the phage and scFv ELISAs. These results confirm that our AFM based biopanning process is a very powerful technique that can be used to generate reagents that selectively bind disease-specific protein variants directly from complex sources.
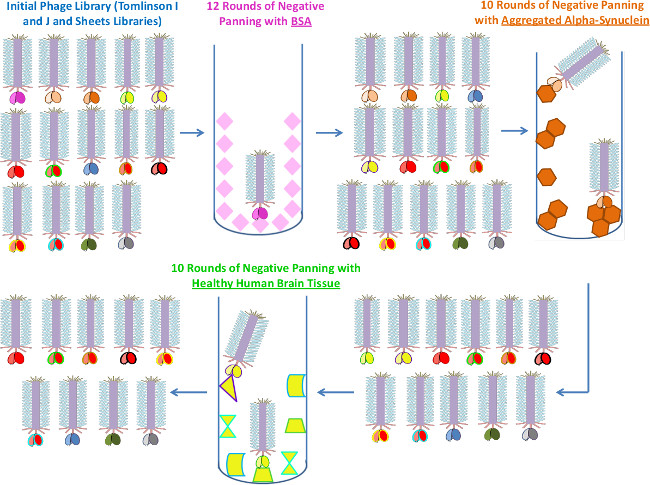
Figure 1. Negative Biopanning Process Utilizing Immunotubes. Schematic demonstrating the negative biopanning process. Phages that are reactive to proteins such as BSA, aggregated alpha-synuclein and healthy brain tissue are removed using immunotubes. Please click here to view a larger version of this figure.

Figure 2. Confirmation of Negative Panning Results. AFM imaging is used to monitor the negative panning steps against the various targets to ensure all reactive phages are removed. Here we show some of the AFM results demonstrating the level of phage binding before and after the negative panning. (A) Phage binding can be detected to aggregated alpha-synuclein particles after the first round of negative panning, but (B) no phage are visible after 8 rounds of negative panning. (C) After the first round of negative panning against healthy tissue phage binding is evident, but (D) no binding is detected after 10 rounds of negative panning. All images are 5 μm. The large white structures on the AFM images are usually due to salts present in the buffers or residual PEG from the PEG precipitation step during phage production. Please click here to view a larger version of this figure.
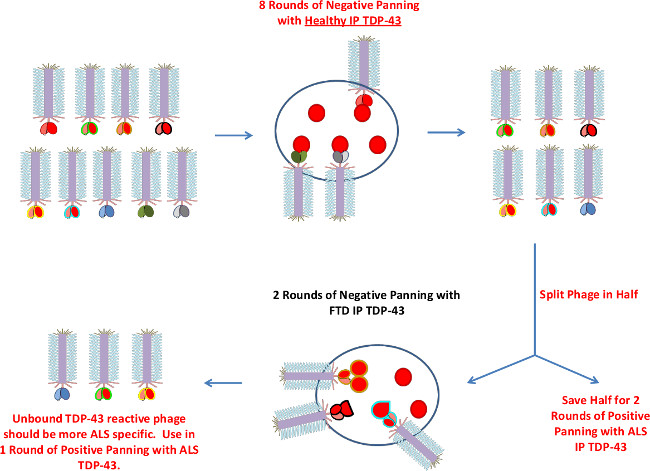
Figure 3. Negative Biopanning Process Utilizing Mica. Schematic demonstrating additional negative biopanning using mica. Due to limited sample mica surface is employed to first eliminate binders to healthy and then FTD immunoprecipitated TDP-43. Before proceeding to the two rounds of negative panning against FTD TDP-43, the phage is split in half (in the event of unsuccessful isolation of ALS TDP-43 exclusive phages during the positive panning phase). Please click here to view a larger version of this figure.
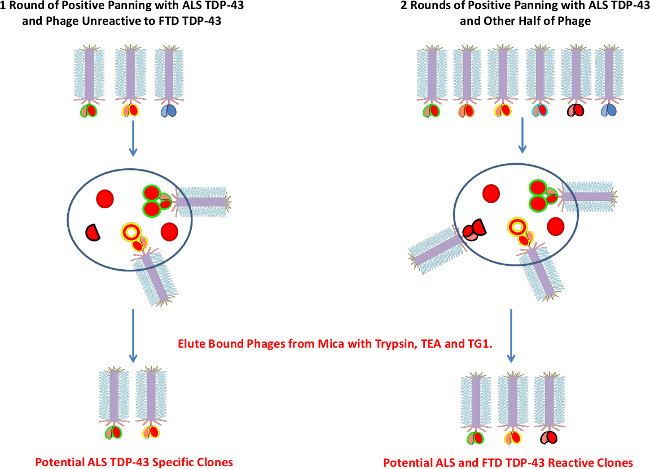
Figure 4. ALS Positive Biopanning Process. Schematic demonstrating the ALS positive biopanning process. The unbound phages after the negative panning against FTD TDP-43 are used in a round of positive panning against ALS TDP-43 to elute more ALS TDP-43 specific clones. Also, two rounds of positive panning against ALS TDP-43 are carried out using mica surface and the unbound phages after negative panning against healthy immunoprecipitated TDP-43 (the bound phages are eluted since these phages should not bind healthy TDP-43, however some may be cross-reactive with both ALS and FTD). Three elution methods are used (trypsin, TEA and TG1 cells) to ensure all bound phages are removed. Please click here to view a larger version of this figure.
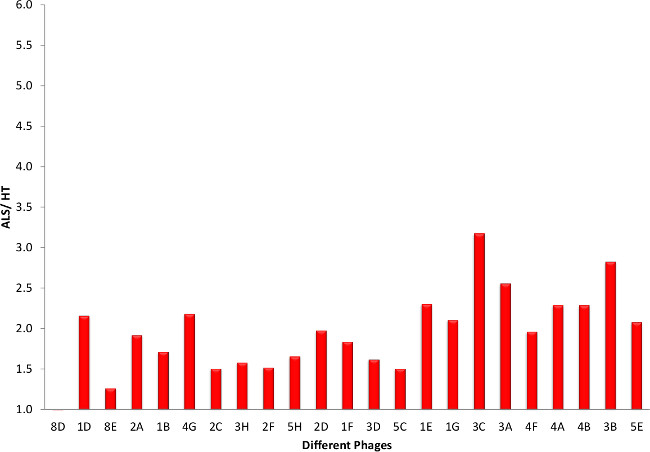
Figure 5. Phage ELISA Screening of Potential ALS Clones (ALS versus HT). Using homogenized human ALS, FTD and healthy brain tissue (HT) samples we performed indirect ELISA using phage produced from the different clones. Results are represented as the ratio to healthy tissue samples. Results showed that some clones distinguish between TDP-43 found in ALS patients and those in healthy. The ALS, FTD and HT tissues are a mix of brain samples (motor cortex) from three individuals. Please click here to view a larger version of this figure.
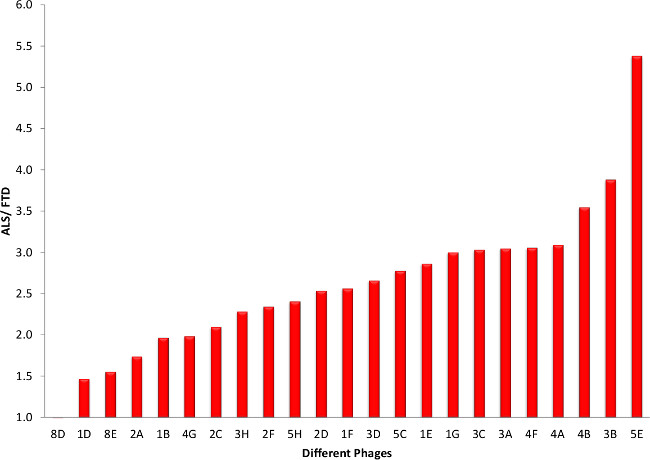
Figure 6. Phage ELISA Screening of Potential ALS Clones (ALS versus FTD). Here we show the phage ELISA results of ALS versus FTD patients. Most of the clones have a preference for ALS over FTD. Please click here to view a larger version of this figure.
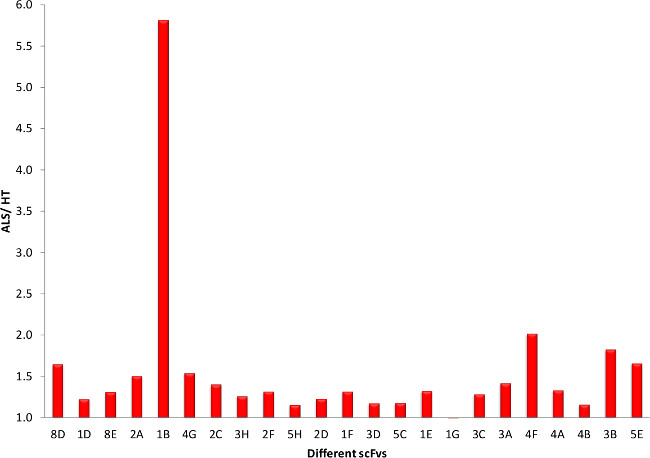
Figure 7. scFv ELISA Screening of Potential ALS Clones (ALS versus HT). Using the scFvs produced from each clone we can observe clones that have a preference for the TDP-43 from ALS patients over healthy. Please click here to view a larger version of this figure.
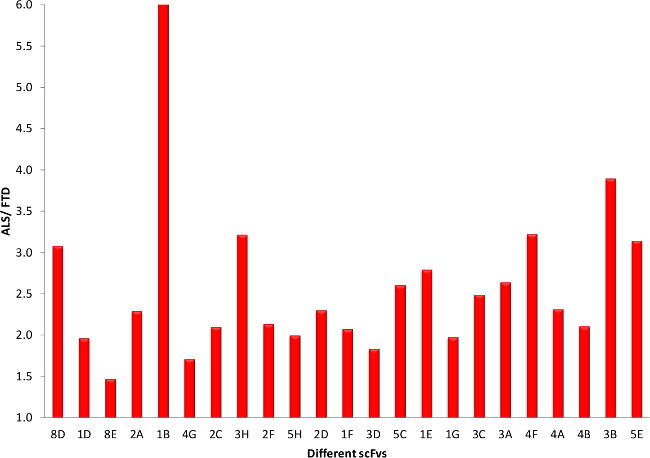
Figure 8. scFv ELISA Screening of Potential ALS Clones (ALS versus FTD). The success of our panning process is further demonstrated when comparing ALS to FTD for the different scFvs in the indirect ELISAs. Please click here to view a larger version of this figure.
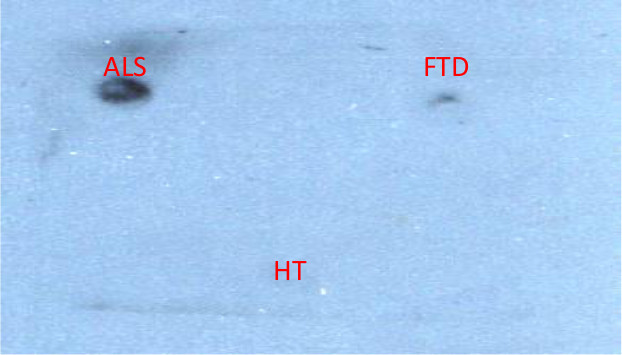
Figure 9. Dot Blot Analysis of Potential ALS Clones. Using dot blot instead of ELISA is another technique to show specificity of the clones for ALS over FTD or HT. Clone 2A is shown. Please click here to view a larger version of this figure.
