This manuscript describes the process required, shown in schematic form in Figure 1, to culture 3D models of the human esophageal epithelium successfully. To confirm the suitability of the model as an experimental platform histological and immunohistochemical studies have been undertaken comparing the cultured tissues with normal human esophageal squamous mucosa.
Histological assessment of the epithelium produced by the method described shows a mature, multi-layered, stratified squamous epithelium (Figure 2B) which is comparable to that observed with the normal human esophagus (Figure 2A), albeit thinner (5 to 10 layers of cells compared with 10 to 20 for the normal esophagus), with the cells becoming progressively flatter and ultimately anuclear as they migrate towards the surface.
Immunohistochemical characterization of key markers of proliferation and differentiation demonstrate that the microanatomy of the model epithelium is similar to the normal human esophageal epithelium. Comparable Ki67 expression is observed in both the native esophagus and the model epithelium, with staining restricted to a subset of cells within the basal and immediately suprabasal layers (Figures 3A and 3B). This is analogous to studies which report that less than 10% of cells generally show expression of the proliferation marker, Ki67, in the normal esophageal epithelium18. CK4 is normally expressed in stratified and columnar epithelia but is generally absent from the basal layers while CK14 is only positive in the basal layer19. In both the normal and model esophageal epithelia, CK14 is observed in all cells in the basal layer (Figures 3D and 3E) while CK4 is observed throughout the epithelium except for the two most basal layers (Figures 3G and 3H). Involucrin expression is an early marker of differentiation, expressed in the suprabasal layers of the epithelium20. Again both the normal human esophageal tissue and the model epithelium show staining that reflects this (Figures 3J and 3K).
Attempts to replace the primary esophageal epithelial cells with immortalized esophageal epithelial cells, such as Het-1A, were less successful and did not produce a valid model of the normal esophageal epithelium. A multi-layered epithelium was formed; however the proliferative marker Ki67 was detected throughout the epithelium (Figure 3C) with no expression detected for any markers of differentiation (Figures 3F, 3I and 3L), indicating that the Het-1A cells produced a hyperproliferative epithelium with no evidence of normal stratification or maturation.
However, the model has been successfully modified to incorporate tumor cells by the replacement of primary epithelial cells with either esophageal adenocarcinoma (OE33) or squamous carcinoma (OE21) cells. This demonstrates the flexibility of the model, enabling its use in investigating a range of esophageal disorders at different stages of progression through the inclusion of a number of different cell lines. It can be seen that there is a marked difference in the responses from these two cell lines. OE21 squamous carcinoma cells produces an epithelium visible on the construct as a defined yellow region (Figure 4A) with large clefts within the epithelium (Figure 4B), likely to reflect dysfunctional cell adhesion molecules. Including OE33 adenocarcinoma cells within the model results in a large amount of scaffold degradation, visible by eye as a thinning of the scaffold after 2 weeks growth at the air/liquid interface (Figure 4C) and confirmed in H+E analysis as an obvious reduction in the thickness of the scaffold in the region below the cells (Figure 4D). This is a likely to be a result of an interaction between the tumor cells and fibroblasts, since the scaffold degeneration is not observed in the absence of fibroblasts (Figures 4E and 4F). We have observed similar impacts on tumor invasion in the presence and absence of fibroblasts in equivalent melanoma models21.
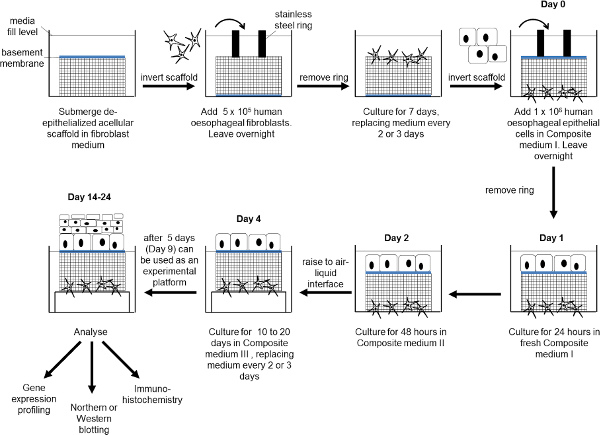
Figure 1: Schematic diagram showing the production of the esophageal mucosa model. Human esophageal fibroblasts are seeded onto the submucosal surface of the scaffold and cultured for 7 days. The scaffold is inverted, human esophageal epithelial cells added and cultured submerged for 4 days. The construct is raised to an air-liquid interface for between 10 and 20 days. At the end of the experiment the construct can undergo further analysis as required. Please click here to view a larger version of the figure.
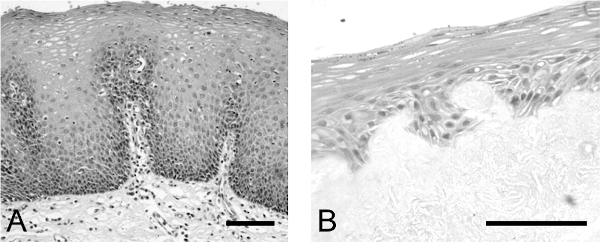
Figure 2: Comparison of epithelium produced in esophageal model with normal human esophageal epithelium. H+E analysis of (A) normal human esophageal epithelium and (B) esophageal epithelium formed in the model of the human esophageal mucosa. Scale bar is 500 µm.Please click here to view a larger version of the figure.
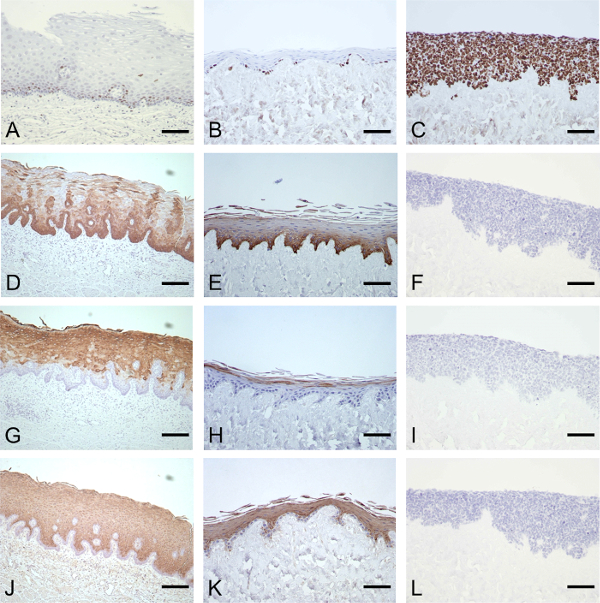
Figure 3: Immunohistochemical characterization of the esophageal epithelium produced in the model and comparison with the normal human esophagus. IHC analysis of normal esophagus (column 1) and the model esophagus produced using primary human esophageal epithelial cells (column 2) or immortalized esophageal epithelial cells (column 3). Characterization was by Ki67 (A–C), CK14 (D, E), CK4 (G, H) and involucrin staining (J, K). Scale bar is 200 µm. (This figure has been modified7.) Please click here to view a larger version of the figure.
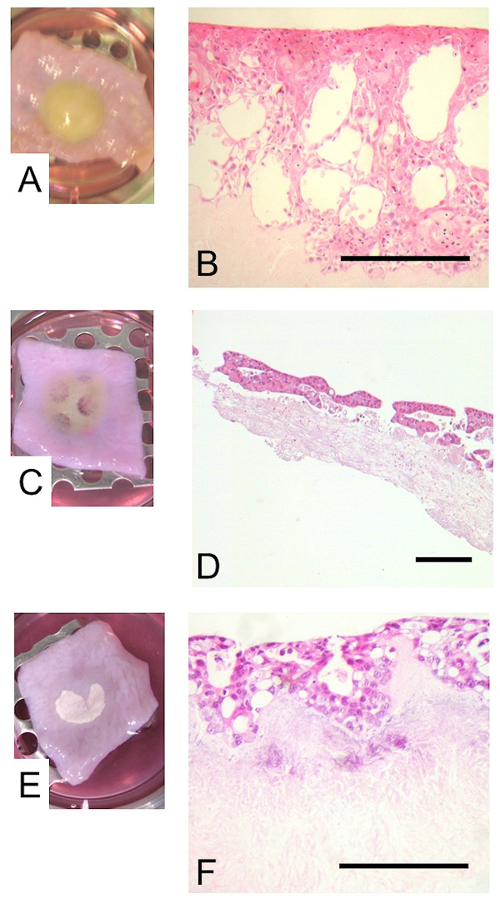
Figure 4: The inclusion of tumor cell lines within the esophageal model. The primary epithelial cells were replaced by the squamous carcinoma cell line, OE21, or the adenocarcinoma cell line, OE33. Images show the construct with the OE21 cells (A) or OE33 cells either in conjunction with (C) or in the absence of (E) fibroblasts, prior to fixing at the end of the culture period. The epithelia including the OE21 (B) or OE33 cells with (D) or without (F) fibroblasts are visualized by H+E staining. Scale bar is 500 µm. Please click here to view a larger version of the figure.
