Т-лимфоциты являются филиал адаптивной иммунной системы характеризуется способностью эффективно распознавать пептид антигена (АГ), связанного с главным комплексом гистосовместимости (МНС) молекул через их Т-клеточных рецепторов (TCR) 1. Наивные лимфоциты мигрируют конструктивно и сканирования »профессиональные Ag представляющих клеток (АПК»; например, дендритные клетки) в лимфатических узлах, в то время как память / эффекторные Т-клетки должны эффективно обследовать чрезвычайно широкий спектр БТР и потенциальных целевых клеток в периферических тканях.
В следующем мин первоначального признания родственных Ag на БТР, лимфоциты арестовать их миграции и начинают формировать специализированные интимную интерфейс клетка-клетка называется "иммунологическое синапсов» (IS). Устойчивый (т.е. 30-60 мин) контакты должны усиливать и поддерживать сигнализации 2-7. Новые исследования определить, что в есть, это непрерывное образование и быстрое гemodeling дискретных субклеточном сигнализации микро-кластеров (т.е., содержащих МНС / Ag-TCR, F-актин, адгезию и сигнальные молекулы), которые определяют силу и качество в результате иммунных ответов 2-7. Тем не менее, динамические данные и механизм регулирования этого процесса полностью понял 8,9. Это связано в основном с техническими проблемами, связанными с неправильной топологии поверхностей APC и плохо контролируемой ориентации плоскостей взаимодействия клетка-клетка, вопросов, которые глубоко ограничивают необходимую пространственно-временной обработки изображений подходов 8-10 (Figure1A).
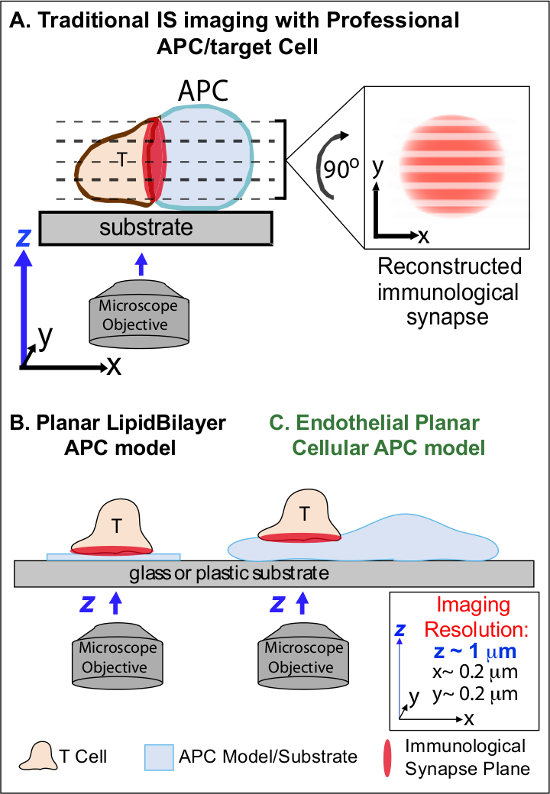
Рисунок 1. Физиологическая Planar сотовый APC Модель изображений иммунологии Synapse Dynamics. Схема иллюстрирует традиционный изображений иммунологической синапса между Т-клетки и профес NAL APC (А) и Т-клеток и традиционный плоский липидов модель бислой APC (В) по сравнению с этой новой модели эндотелиальной плоской APC (C). Профессиональные БТР обеспечить физиологические иммунологические синапсы, но предлагают мало ориентированный интерфейс клетка-клетка (т.е., по отношению к оптимальной плоскости изображения ху, разрешение ~ 0,2 мкм), что резко компрометирует пространственное (г визуализации самолет разрешения ~ 1 мкм) и временная (т.е. в связи с необходимостью многократно сканировать всех плоскостях изображений г) разрешение изображения. Двухслойные модели имеют плоскую топологию, что обеспечивает оптимальную пространственно-временной изображения разрешением, но также сильно упрощенной, не физиологические и жесткой. Это эндотелия модель сотового сочетает в себе плоскую топологию липидного бислоя с физиологической подложки классического APC, чтобы доставить оптимального пространственное и временное разрешение изображений в физиологическом обстановке.м / файлы / ftp_upload / 53288 / 53288fig1large.jpg "целевых =" _blank "> Нажмите здесь, чтобы посмотреть большую версию этой фигуры.
Предыдущие исследования частично обойдена эти препятствия путем разработки моделей плоскую подложку (т.е. липидных бислоев и поверхностей покрытых антителами), которые обеспечивают оптимальную пространственно-временной разрешение (т.е. с помощью фиксации поверхность активации Т-клеток в единый план, параллельной оптимальной визуализации ху Плоскость) 11-15 (Фигура 1В). Эти модели способствовали важные идеи в субклеточных / молекулярной динамики, которые контролируют антигенный сигнализации в Т-клетках, в том числе открытие динамического актина / TCR сигнализации микро-кластеров 7,11-14. Тем не менее, такие модели изначально упрощать, а также жесткой (исключающее развитие / изучение 3-мерных топологических особенностей) (рис 1B). Таким образом, остается неясным, как связать такие выводы PHYsiologic клетка-клетка иммунный надзор.
Хотя до сих пор изучено недостаточно, сосудистой и лимфатические эндотелиальные клетки возникают, как большой (т.е. больше, чем в цифрах всех профессиональных БТР, на ~ 1000 раз) периферической отсеке "полу-профессиональной" БТР 16-18. Эти клетки экспрессируют MHC-I-, MHC-II- и множество молекул со-стимуляторов (например, CD40, LFA3, ICOSL, 4-1ВВ, OX40L, TL1A, PD-L1, но не CD80 и CD86) и стратегически расположенный на границе кровь-ткань, где они служат специализированные функции дозорных 16-18. Предыдущие исследования показали, что эндотелиальные клетки могут эффективно повторно стимулировать эффекторной / память, но не наивным, Т-клетки 19-25. Таким образом, эндотелиальные клетки, вероятно, играют уникальную роль в эффекторной APC фазе адаптивных иммунных реакций в пределах периферических тканях, таких как локального воздействия на активацию Т-клеток, дифференцировки, памяти и толерантности 16,17,26. Крически, при выращивании в пробирке, эндотелиальные клетки образуют практически плоские поверхности клеток и легко transfectable (например, с флуоресцентным белком журналистами). Эти особенности идеально подходят для высокой пространственно-временной разрешающей способностью топологических динамики при межклеточных взаимодействий 19,27. Таким образом, эндотелиальные клетки может служить физиологический "плоской сотовой APC» модели отчетливо подходит для изучения субклеточных / молекулярные механизмы ремоделирования, которые управляют распознавания антигенов и регулируют ответов (рис 1в) 19,20.
Ранее установлено, комплементарные методы визуализации (в том числе трансфекции эндотелии клеток с флуоресцентными производителей белка мембраны плазмы и цитозоле) для изучения деталей лейкоцитов-эндотелиальной взаимодействия при адгезии и трансэндотелиальной миграции 27, показали, что лейкоциты активно зондировать поверхность эндотелии динамического вставкад втягивание субмикронных масштабах, актин-богатых цилиндрические выступы (~ 200-1000 нм в диаметре и глубиной) называется invadosome, как выступы (то есть, "ILPs ') 27,28. Эти подходы изображений были расширены наряду с созданием протоколов, чтобы воспользоваться функцией эндотелия APC развивать первые методы пространственно-временной высокой разрешающей способностью в Т-клеток-эндотелиальной иммунологической синапса Как сообщалось 19,20 и далее описанных здесь. Центральный вывод, производный от этого романа плоской модели APC сотовой том, что Т-клеточные ILPs функционировать как в поощрении начального обнаружения Ag и в поддержании последующее сигнализации. В самом деле, массивы нескольких ILPs (которые были стабилизированы и начисленных в ответ на начальный поток кальция) Показать обогащение TCR и молекул наводящий активного сигнального такой ПКС-Q, ZAP-70, фосфотирозина и HS1. Таким образом, ILPs-видимому, представляют собой трехмерную физиологическую эквивалентную ТКР сигнализации микрокластеры видели в плоских моделей двухслойных. Этот подход, таким образом, чутко показывает / отчеты молекулярные и архитектурные (и подразумеваемые биомеханические) динамика не иначе обнаруживается.
Метод, описанный в данном документе должны быть полезными для преодоления разрыва между профессиональной APC и искусственных моделей APC субстрата для того, чтобы укрепить нашу способность допросить основные механизмы адаптивного иммунного ответа. В то время как здесь акцент делается на активацию CD4 + Th1-типа эффектора / ячейки памяти, это основной подход можно легко модифицировать для изучения широкий спектр типов Т-клеток и AGS, как описано ниже.
