לימפוציטים מסוג T הוא סניף של המערכת החיסונית אדפטיבית המאופיינת ביכולת לזהות ביעילות אנטיגן פפטיד (Ag) חייבת מורכב histocompatibility גדול (MHC) מולקולות באמצעות קולטנים של תאי T (TCRs) 1. לימפוציטים נאיביים constitutively להעביר ולסרוק תאים מקצועיים Ag הצגה '(נגמ"שים; למשל, תאים דנדריטים) בתוך בלוטות הלימפה, בעוד תאי הזיכרון / T מפעיל צריכים לסקור מגוון רחב מאוד של נגמ"שים ותאי יעד פוטנציאליים בתוך רקמות היקפי ביעילות.
בדקות הבאות הכרה ראשונית של Ag מקור על APC, ימפוציטים לעצור הגירתם ומתחיל להיווצר ממשק תאי תאים אינטימי מיוחד "סינפסה החיסונית 'כינתה (IS). ספג (כלומר, 30-60 דקות) היא קשר נדרש להגביר ולקיים איתות 2-7. מחקרים המתעוררים לזהות שבתוך הוא, זה הוא ההיווצרות רציפה ומהיר remodeling של מיקרו-אשכולות בדידים איתות תת-תאי (כלומר, מכילים MHC / Ag-TCR, F- אקטין, הדבקה ומולקולות איתות) הקובעים את החוזק והאיכות של תגובות חיסוני וכתוצאה מכך 2-7. עם זאת, פרטים דינמיים ומנגנון רגולציה של תהליך זה באופן חלקי הבינו 8,9. זה נובע במידה רבה מאתגרים טכניים הקשורים בטופולוגיות סדירות של משטחי APC ונטייה גרועים מבוקרת של מטוסי האינטראקציה תאי תאים, נושאים שעמוקים להגביל את ההדמיה spatiotemporal הנדרשת גישות 8-10 (Figure1A).
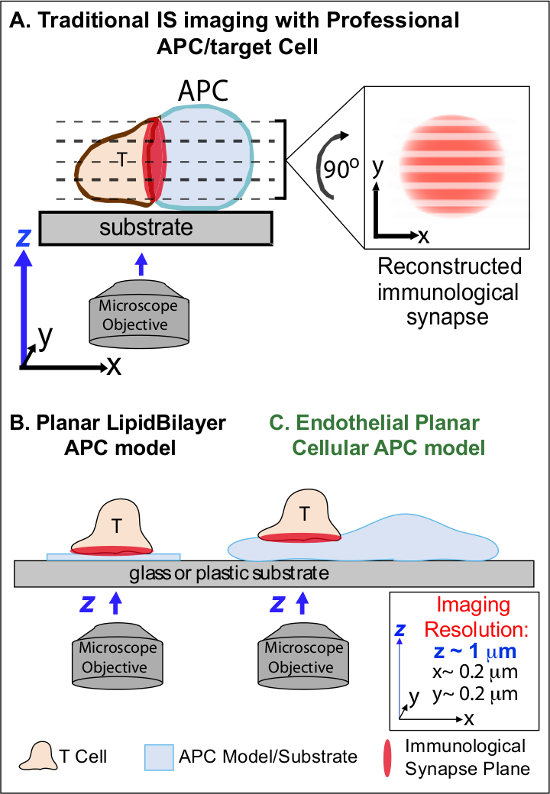
איור 1. דגם APC התא פיסיולוגי מישורי הדמיה החיסוני סינפסה Dynamics. סכמטי ממחיש הדמיה מסורתית של סינפסה החיסונית בין תאי T וprofessio תא סופי APC () ו- T ומודל מסורתי שומנים מישוריים APC bilayer (ב ') בהשוואה למודל זה רומן אנדותל APC מישוריים (C). נגמ"שים מקצועיים לספק סינפסות החיסונית פיזיולוגית אבל מציעים ממשק מונחה גרוע תא-תא (כלומר, ביחס למישור ההדמיה XY האופטימלי; רזולוציה ~ 0.2 מיקרומטר), אשר באופן דרמטי פוגע מרחבית (רזולוציה ~ מיקרומטר מטוס ההדמיה z 1) ובזמן (כלומר, בשל הצורך לסרוק שוב ושוב בכל מטוסי ההדמיה z) רזולוציה של הדמיה. יש דגמי bilayer טופולוגיה מישוריים המספקת רזולוציה ההדמיה spatiotemporal אופטימלית, אבל גם פשוטים ביותר, שאינם פיסיולוגי ונוקשה. מודל תא האנדותל זה משלב את הטופולוגיה מישוריים של bilayers שומנים עם המצע הפיזיולוגי של APC קלאסי כדי לספק רזולוציה הדמיה מרחב ובזמן אופטימלית בסביבה פיזיולוגית.מ '/ קבצים / ftp_upload / 53,288 53288fig1large.jpg "target =" / _ blank "> לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
העבודה קודמת עקפה באופן חלקי על המכשולים הללו על ידי פיתוח מודלים מצע מישוריים (כלומר, bilayers שומנים ומשטחים מצופים נוגדנים) המספקים רזולוציה spatiotemporal אופטימלית (כלומר, באמצעות תיקון משטח הפעלת תא T לתכנית אחת שמקבילה להדמית XY האופטימלית מטוס) 11-15 (1B איור). מודלים אלה הקלו תובנות חשובות דינמיקת subcellular / מולקולרית השולטות איתות אנטיגני בתאי T, כוללים גילוי אקטין דינמי / TCR מיקרו-אשכולות איתות 7,11-14. עם זאת, מודלים כאלה מטבעם פשטניים, כמו גם נוקשה (מניעת הפיתוח / המחקר של תכונות טופולוגי 3 ממדים) (איור 1). לכן, זה עדיין לא ברור איך להתייחס כגון ממצאים לPHYתאי תאים siologic מעקב חיסוני.
למרות שעדיין understudied, כלי דם ותאי האנדותל הלימפה הם מתעוררים כגדולים (כלומר, גדולים יותר במספרים מכל נגמ"שים המקצועיים, על ידי ~ 1000-פי) תא היקפי של נגמ"שים "חצי מקצועי" 16-18. תאים אלה מבטאים MHC-אני-, MHC-II- ושפע של מולקולות שיתוף ממריץ (למשל, CD40, LFA3, ICOSL, 4-1BB, OX40L, TL1A, PD-L1, אבל לא CD80 וCD86) ואסטרטגיים ממוקם בממשק שבין דם לרקמה שבו הם משרתים פונקציות זקיף מיוחדות 16-18. מחקרים קודמים הראו שתאי האנדותל יכולים למעשה מחדש לעורר מפעיל / זיכרון, אבל לא נאיבי, תאי T 19-25. כך, תאי האנדותל עשויים לשחק תפקידי APC ייחודיים בשלב מפעיל של תגובות חיסוניים אדפטיבית בתוך הרקמות ההיקפית, כגון השפעה מקומית על הפעלת תא T, בידול, זיכרון וסובלנות 16,17,26. CRItically, כאשר גדלו במבחנה, תאי האנדותל יוצרים משטחי תא כמעט מישורי והם בקלות transfectable (למשל, עם כתבי חלבון פלואורסצנטי). תכונות אלה הן אידיאליים עבור הדמיה גבוהה spatiotemporal רזולוציה של דינמיקה טופולוגית במהלך אינטראקציות תא-תא 19,27. כך תאי האנדותל עשויים לשמש כמודל 'מישוריים סלולארי APC' פיסיולוגי מובהק מתאים ללימוד מנגנוני שיפוץ subcellular / מולקולריים המניעה את הכרת אנטיגן ולווסת תגובות (איור 1 ג) 19,20.
בעבר הוקם טכניקות הדמיה משלימות (כולל transfection של תאי endothelia עם מקבלי חלבון פלואורסצנטי של קרום הפלזמה וcytosol) ללומד את הפרטים של אינטראקציה ליקוציט-האנדותל בהידבקות וההגירה Transendothelial 27, הראה כי לויקוציטים באופן פעיל לחקור את פני השטח של האנדותל ידי דינמי הכנסהההכחשה ד של תת-מיקרון בקנה מידה, בליטות גליליות אקטין עשיר (~ 200-1,000 ננומטר בקוטר ועומק) כינו בליטות כמו invadosome (כלומר, 'ILPs') 27,28. הדמיה גישות אלה הורחבו עוד יותר יחד עם יצירת הפרוטוקולים לנצל פונקצית APC אנדותל לפתח השיטות הראשונות להדמיה גבוהה spatiotemporal רזולוציה של סינפסה החיסונית תא אנדותל T כ19,20 דיווח ועוד לתאר זאת. ממצא מרכזי הנגזר ממודל APC סלולארי זה מישוריים רומן הוא שILPs תא T לתפקד גם בקידום זיהוי Ag ראשוני ובשמירת איתות שלאחר מכן. ואכן, מערכים של ILPs מרובה (שהתייצבו ונצבר בתגובה הראשונית שטף סידן) להעשרת תכנית בTCR ומולקולות רמיזות של PKC-Q איתות פעילה כגון, ZAP-70, phosphotyrosine וHS1. לכן, ILPs נראה לייצג מקביל פיסיולוגי תלת-ממדית למייקרו-TCR האיתותאשכולות ראו בדגמי bilayer מישוריים. גישה זו, ובכך, ברגישות מגלה / דינמיקה מולקולרית ואדריכלית (ומשתמע ביו-מכאנית) דיווחים לא אחרת לזיהוי.
השיטה המתוארת במסמך זה צריכה להיות שימושי לגישור על הפער בין APC המקצועי ודגמי מצע APC מלאכותיים כדי לשפר את היכולת שלנו לחקור את המנגנונים בסיסיים של תגובות חיסוניים אדפטיבית. ואילו כאן הדגש הוא על ההפעלה של מפעיל CD4 + סוג Th1 / תא זיכרון, גישה בסיסית זו ניתן לשנות בקלות ללמוד מגוון רחב של סוגי תאי T וAGS, כמפורט להלן.
