Los linfocitos T son una rama del sistema inmune adaptativo que se caracteriza por la capacidad de reconocer eficientemente péptido antígeno (Ag) unida a complejo mayor de histocompatibilidad (MHC) moléculas a través de sus receptores de células T (TCR) 1. Linfocitos ingenuos constitutivamente migran y escanear 'Ag células presentadoras profesionales' (APC; por ejemplo, células dendríticas) en los ganglios linfáticos, mientras que las células de memoria / T efectoras necesitan para estudiar con eficacia una muy amplia gama de vehículos blindados y potenciales células diana dentro de los tejidos periféricos.
En el min tras el reconocimiento inicial de cognado Ag en un APC, linfocitos detener su migración y comienzan a formar una interfaz célula-célula íntima especializado denominado 'sinapsis inmunológica "(IS). Sostenida (es decir, 30 a 60 min) ES se requieren contactos para ampliar y mantener la señalización 2-7. Estudios emergentes identificar que dentro del IS, que es la formación continua y rápida remodeling de señalización subcelular micro-grupos discretos (es decir, que contienen MHC / Ag-TCR, F-actina, la adhesión y moléculas de señalización) que determinan la fuerza y la calidad de la que resulta la respuesta inmune 2-7. Sin embargo, los detalles dinámicos y mecanismo de regulación de este proceso se conocen por completo 8,9. Esto se debe en gran parte de los desafíos técnicos asociados con topologías irregulares de las superficies de APC y la orientación de un mal control de los planos de interacción célula-célula, las cuestiones que limitan profundamente la imagen espacio-temporal necesaria acerca 10.08 (Figure1A).
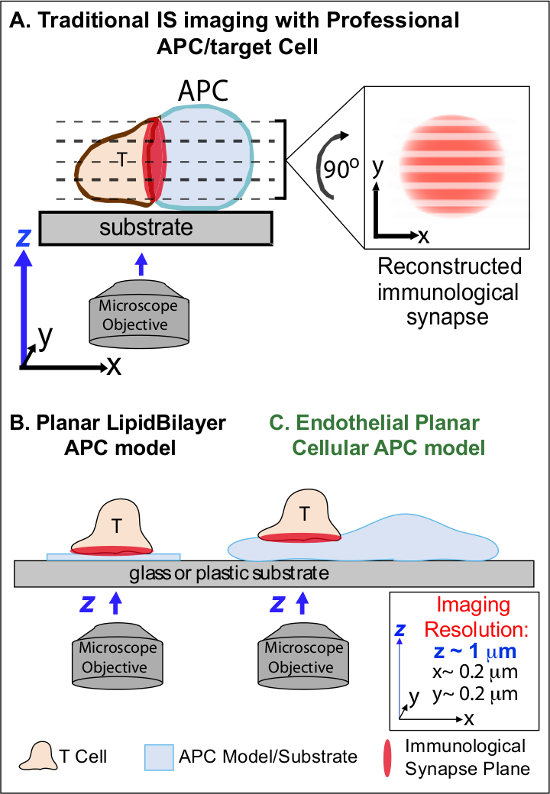
Figura 1. Un fisiológica planar Modelo APC Cell for Imaging inmunológica Synapse Dynamics. El esquema ilustra la imagen tradicional de la sinapsis inmunológica entre una célula T y un professio nal de células APC (A) y T y un modelo tradicional de lípidos planar APC bicapa (B) en comparación con este novedoso modelo de APC planar endotelial (C). APCs profesionales proporcionan sinapsis inmunológicas fisiológicas pero ofrecen interfaz mal orientada célula-célula (es decir, con respecto al plano óptima de imagen xy; resolución ~ 0,2 micras), lo que compromete drásticamente espacial (z plano de la imagen de resolución ~ 1 m) y temporal (es decir, debido a la necesidad de escanear repetidamente a través de todos los planos de formación de imágenes z) resolución de la imagen. Modelos bicapa tienen una topología plana que ofrece óptima de imágenes de resolución espacio-temporal, pero también son muy simplificada, no fisiológico y rígido. Este modelo de células endoteliales combina la topología planar de bicapas lipídicas con el sustrato fisiológico de un APC clásico para entregar óptima de imágenes de resolución espacial y temporal en un entorno fisiológico.m / files / ftp_upload / 53288 / 53288fig1large.jpg "target =" _ blank "> Haga clic aquí para ver una versión más grande de esta figura.
El trabajo previo ha eludido parcialmente estos obstáculos mediante el desarrollo de modelos sustrato plano (es decir, bicapas lipídicas y las superficies recubiertas de anticuerpo) que proporcionan resolución espacio-temporal óptima (es decir, a través de la fijación de la superficie de activación de las células T en un único plan que es paralela a la proyección de imagen óptima xy avión) 11 a 15 (Figura 1B). Estos modelos han facilitado importantes conocimientos sobre la dinámica subcelulares / moleculares que controlan la señalización antigénico en las células T, incluyendo el descubrimiento de actina dinámica / TCR señalización micro-clusters 7,11-14. Sin embargo, tales modelos están inherentemente simplificaron, así como rígido (que impide el desarrollo / estudio de características topológicas 3 dimensiones) (Figura 1B). Por lo tanto, sigue siendo incierto cómo relacionar estos hallazgos a physiologic célula-célula vigilancia inmune.
Aunque todavía poco estudiado, vascular y las células endoteliales linfáticas están emergiendo como un grande (es decir, mayor en número que todos los vehículos blindados profesionales, por ~ 1000 veces) compartimento periférico de "semi-profesional" APC 16-18. Estas células expresan MHC-I, MHC-II- y una multitud de moléculas co-estimuladoras (por ejemplo, CD40, LFA3, ICOSL, el 4-1BB, OX40L, TL1A, PD-L1; pero no CD80 y CD86) y son estratégicamente posicionado en la interfase sangre-tejido donde sirven funciones especializadas centinela 16-18. Estudios previos demostraron que las células endoteliales pueden efectivamente re-estimular efectoras / memoria, pero no ingenuo, las células T 19-25. Así, las células endoteliales es probable que desempeñen un papel único de APC en la fase efectora de la respuesta inmune adaptativa dentro de los tejidos periféricos, tales como la influencia local sobre la activación de células T, la diferenciación, la memoria y la tolerancia 16-17,26. Cricamente, cuando se cultivan in vitro, las células endoteliales forman superficies celulares prácticamente planas y están fácilmente transfectable (por ejemplo, con los reporteros fluorescentes de proteínas). Estas características son ideales para imágenes de alta resolución espacio-temporal de la dinámica topológicos durante las interacciones célula-célula 19,27. Por lo tanto las células endoteliales podrían servir como un 'celular plana APC modelo fisiológica claramente adecuado para el estudio de los mecanismos de remodelación subcelulares / molecular que impulsan el reconocimiento de antígenos y regulan las respuestas (Figura 1C) 19,20.
Previamente establecidos técnicas de imagen complementarios (incluyendo la transfección de las células endoteliales con los fabricantes de proteínas fluorescentes de la membrana plasmática y citosol) para estudiar los detalles de la interacción de leucocitos endotelial durante la adhesión y migración transendotelial 27, mostraron que los leucocitos sondean activamente la superficie del endotelio por Dynamic una inserciónd retracción de la sub-escala micrométrica, protuberancias cilíndricas de actina-ricos (~ 200-1.000 nm de diámetro y profundidad) denomina-invadosome como salientes (es decir, 'ILPS') 27,28. Estos enfoques de imagen se han ampliado aún más, junto con la creación de protocolos para tomar ventaja de la función endotelial de APC para desarrollar los primeros métodos de alta resolución de imagen espacio-temporal de la célula endotelial sinapsis inmunológica T según lo informado 19,20 y describir con más detalle en el presente documento. Un hallazgo central derivado de esta novela plana celular modelo de APC es que programas de vida independiente de células T funcionan tanto en la promoción de la detección inicial Ag y en el mantenimiento de la señalización posterior. De hecho, las matrices de múltiples programas de vida independiente (que fueron estabilizados y devengados en respuesta a sus iniciales en el flujo de calcio) muestran un enriquecimiento en TCR y moléculas sugestivos de señalización activa tal PKC-Q, ZAP-70, fosfotirosina y HS1. Por lo tanto, programas de vida independiente parecen representar un equivalente fisiológico tridimensional a la micro TCR de señalizaciónracimos visto en los modelos de dos capas planas. Este enfoque, por lo tanto, revela sensibilidad / informes dinámica molecular y arquitectónicos (y biomecánico implícitas) de otro modo no detectable.
El método descrito en este documento deberá ser útil para reducir la brecha entre los modelos de sustratos artificiales APC APC profesional y con el fin de mejorar nuestra capacidad de interrogar a los mecanismos básicos de la respuesta inmune adaptativa. Mientras que aquí la atención se centra en la activación de tipo Th1 CD4 + efectoras / célula de memoria, este enfoque básico puede ser modificado fácilmente para estudiar una amplia gama de tipos de células T y Ags, como se discute a continuación.
