Pupal injection for gene KD yields optimal results when injection is performed during the early pupal stage, when cuticle tanning levels are low (Figure 1A, left, and 1B). Increased tanning and hardening of cuticle, generally after 24 hours, results in increased pupal death following injection (Figure 1A, center and right). The rate of pupal development can vary depending on insectary conditions and animal density24,25; therefore, it is best to assess pigmentation visually.
During the injection process, the capillary needle is inserted into the dorsal cuticle at an angle of approximately 30° in the anterior to posterior direction (Figure 2A). Once the needle is inserted and the dsRNA + 0.01% (w/v) FGD is dispensed, the distribution of dye is evident throughout the hemolymph (Figure 2B).
Assessment of adult emergence for pupae injected with 0.01% (w/v) FGD revealed an average rate of 70% emergence, compared to 96.7% emergence of non-injected controls (Figure 3A). Of note, partial emergence from the pupal case was observed for a large number of non-surviving mosquitoes (Figure 3B). Injected animals exhibit no delays in emergence time (Figure 4A) or biased impact on either gender (Figure 4B). Additional assessment of adult survival carried out up to day 10 post-emergence reveals no evident impact on post-emergence adult survival (Figure 4C).
Validation of KD quality was assessed by the melanotic pseudo-tumor (darkly pigmented tissue cluster) phenotype associated with SRPN2 knockdown21,22 as a positive control for knockdown and the absence of phenotypes associated with dsLacZ injection as a negative control. Adult mosquitoes that emerged were scored at day 8 post-injection (day 6-7 post-emergence). Melanotic pseudo-tumors (Figure 5A and 5B) were observed through the cuticle of 93.5% of the dsSRPN2 vs. 0% of the dsLacZ adult mosquitoes (Figure 6A). Clusters of darkly melanized tissue were identified upon dissection of pigmented patches (Figure 5C). Pseudo-tumors were visible on the adult cuticle as early as day 3 post-emergence and were also present in a subset of dsSRPN2 hemolymph and gut tissues (data not shown). At 5 post-injection (early adult stage), significantly decreased SRPN2 levels in dsSRPN2, but not dsLacZ or non-injected hemolymph protein isolates was observed (Figure 6B).
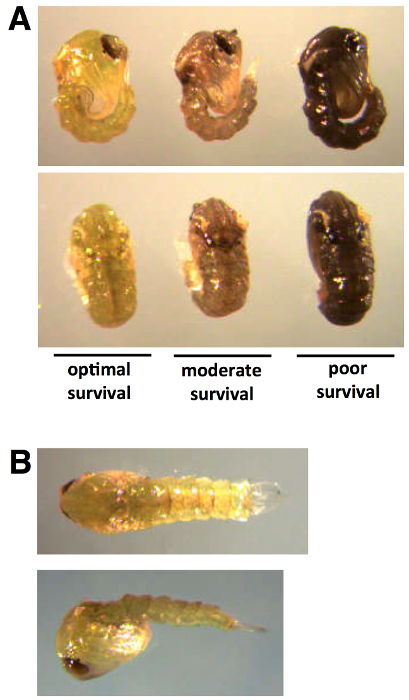
Figure 1: Developmental staging for pupal dsRNA injection. Early pupal injection of dsRNA results in optimal survival and progression into adult stage. Low levels of cuticle pigmentation (A, left, and B) can be observed within the first 0-24 hr following pupation. Tanning of the pupal cuticle preceding injection (A, center and right) results in moderate to poor survival.
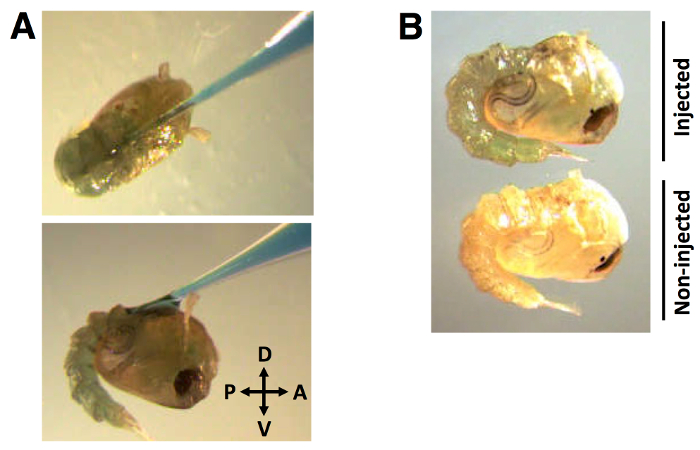
Figure 2: Injection position and distribution of dye-labeled dsRNA. (A) Capillary needle injection of dye-labeled dsRNA into the dorsal cuticle at an angle of approximately 30°, in anterior to posterior direction. (B) The dye is visibly distributed in the pupal hemolymph. dsRNA injection volume of 138 nl, labeled with 0.01% FGD (w/v).
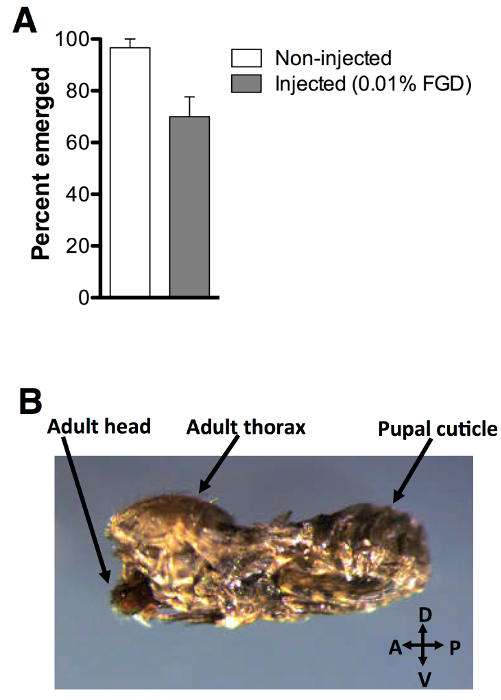
Figure 3: Post-injection adult emergence. (A) 70% of pupae injected with 0.01% FGD (w/v) successfully emerged (n = 60), compared to 96.7% of non-injected controls (n = 60). Three biological replicates were performed. (B) Partial emergence from the pupal case was observed for a large number of non-surviving mosquitoes. Error bars represent the standard error of the mean (SEM).
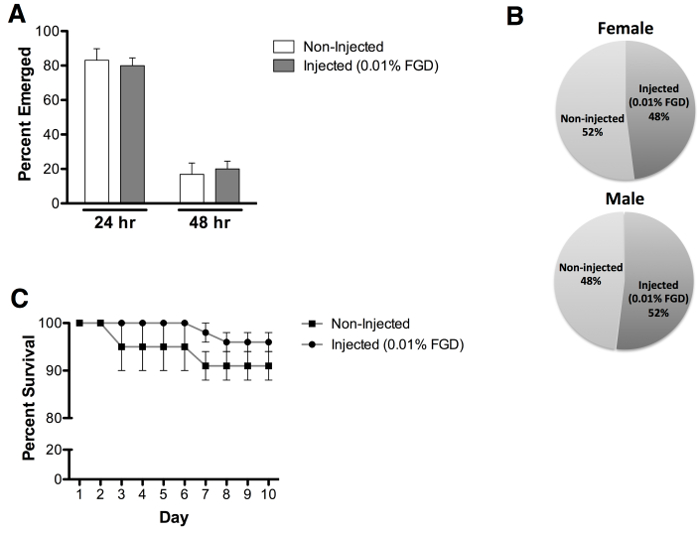
Figure 4: Emergence rate, sex assessment and adult survival. (A) Comparable emergence times were observed following pupal injection with 0.01% FGD (24 hr = 80% and 48 hr = 20%), as compared to non-injected pupae (24 hr = 83% and 48 hr = 17%). (B) Approximately equal male and female adult emergence was observed for 0.01% FGD injected pupae (female = 48% and male = 52%) and non-injected pupae (female = 52% and male = 48%). (C) Survival analysis reveals that injection with 0.01% FGD does not impact adult survival, assessed up to day 10 post-emergence. Results represent data from three independent experiments with 0.01% FGD injected (n = 60) and non-injected (n = 60) pupae (equal numbers of males and females). Error bars represent the standard error of the mean (SEM).

Figure 5: Pseudo-tumor positive control phenotype reflects successful knockdown. Pseudo-tumors were observed on the (A) abdominal and (B) thoracic cuticle of dsSRPN2-injected, but not dsLacZ-injected adult mosquitos at day 8 post-injection. (C) Higher magnification (400X) imaging (a) of cuticle and dissection of pigmented patches (b) reveals clusters of darkly melanized cells.
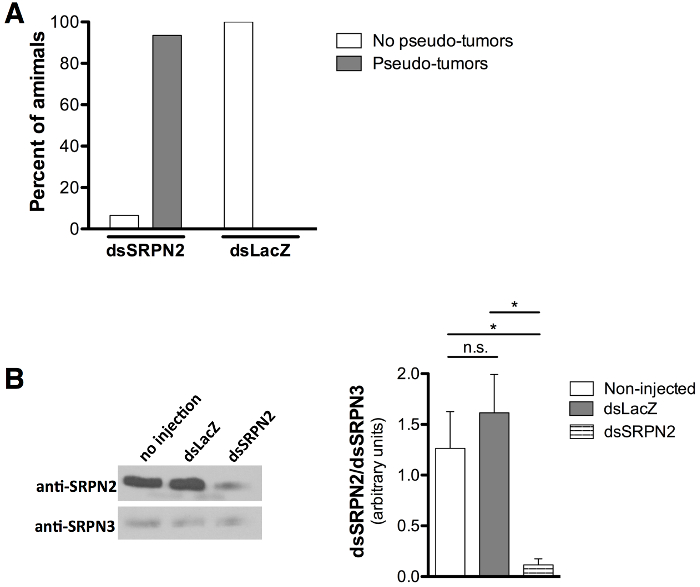
Figure 6: Quantification of pseudo-tumor formation and decreased SRPN2 protein levels. (A) Pupal stage injections result in pseudo-tumor formation in 93.5% of dsSRPN2 adults (n = 21) compared to 0% of dsLacZ controls (n = 19). Results obtained day 8 post-injection. (B) Western blot (left) shows decreased SRPN2 levels in dsSRPN2, but not dsLacZ or non-injected hemolymph protein isolates (day 5 post-injection). Results based on three independent experiments. Anti-SRPN221 and anti-SRPN326 antibody dilutions used were 1:1,000 and 1:2,000, respectively. Goat anti-rabbit IgG-HRP (Product sc-2004, Santa Cruz Biotechnology, Dallas TX) was used at 1:5,000. All protein levels were quantified (right) by band intensity (ImageJ Software, NIH, Bethesda, MD), normalized to SRPN3, and statistically compared by unpaired t test. P <0.05: *, P ≥0.05: n.s. (not significant). Error bars represent the standard error of the mean (SEM).
