There are two key components that allow CaspaseTracker to detect caspase activity in normal healthy cells (Figure 1a). The first of these is a 146 amino acid caspase-cleavable polypeptide modeled after the caspase biosensor Apoliner (Figure 1b).28 This polypeptide is derived from DIAP1 (Drosophila inhibitor of apoptosis) containing a single naturally occurring caspase site that is cleaved during apoptosis typically by the caspase DrICE.42,43 DrICE is equivalent to caspase-3 in mammals and cleaves most known cellular substrates.4,32 Characteristic of caspases, DIAP1 and its derived polypeptide are cleaved after a specific aspartic acid, Asp20 located within the caspase recognition sequence 17-DQVD-20, and mutation of the obligatory Asp20 residue to Ala (DQVA) abolishes cleavage.42,43 Similar to Apoliner,28 this DIAP1 fragment is anchored in the cytoplasm at the plasma membrane via mouse CD8 (alpha chain amino acids 1-220), a commonly used tool in Drosophila. The CD8 membrane anchor prevents any tethered cargo bearing a nuclear translocation sequence (nucGFP in the case of Apoliner) from translocating to the nucleus in the absence of caspase activity.28 The second key component of CaspaseTracker is the Drosophila G-TRACE system in which the yeast transcription factor Gal4 induces the expression of flippase (FLP) recombinase (and simultaneously induces RFP).34 Flippase excises a stop codon leading to permanent expression of nucGFP. To make G-TRACE responsive to caspase activity, it was combined with the Apoliner system by tethering Gal4, which is required to activate G-TRACE, to the plasma membrane-anchored caspase-cleavable DIAP1 fragment of Apoliner (Figure 1a).35
We made additional modifications to the Apoliner component of CaspaseTracker to improve utility. Most important, it is critical that the caspase biosensor itself does not inhibit caspases, potentially preserving cells that would otherwise die. Although the Apoliner transgene did not cause obvious phenotypes in Drosophila28, as would be expected of a potent caspase inhibitor, even the occasional preservation of cells would defeat the purpose of identifying normal cells with caspase activity. However, the DIAP1 fragment in Apoliner contains a motif (135-DICG-138) that was subsequently shown to potently inhibit DrICE, the same caspase that cleaves DIAP1 at Asp20.43 The mechanism of caspase inhibition was revealed by a crystal structure in which DIAP1 Asp135 is bound into the DrICE active site as a caspase substrate mimic (but is not cleaved).43 Biochemical analysis demonstrated that an Arg substitution at Asp135 abolishes DrICE-inhibitory function.43 Therefore, CaspaseTracker was engineered with the same D135R mutation to avoid caspase inhibition (Figure 1b).35 Similar to Apoliner, the naturally occurring DIAP1 Asn-Asn sequence at the de novo N-terminus following cleavage at Asp20 was changed to Gly-Val in CaspaseTracker to prevent degradation of Gal4 by the N-end rule upon caspase cleavage (Figure 1b).28 In addition, we fused a myc-tag to the C-terminus of Gal4 for ready detection of CaspaseTracker expression.35 Of necessity, Apoliner's own RFP and nucGFP cassettes were deleted to make it compatible with G-TRACE, which also contains RFP and nucGFP.28,35
Because Gal4 bears its own nuclear localization signal and potently activates transcription via its target DNA response element (UAS) when fused to other transgenes in Drosophila, presumably only a few molecules of Gal4 released by cytoplasmic caspases are sufficient to turn on RFP expression within a few hours (estimated ≤12 h). Because biosensor activity requires de novo transcription and translation of RFP, it does not report real-time enzymatic activity. Although CaspaseTracker RFP will be significantly degraded likely within a day after caspase activity ceases, nucGFP will be expressed by the ubiquitin promoter for the life of the cell and its progeny.
To first demonstrate that CaspaseTracker is responsive to caspase activation in vivo, adult CaspaseTracker biosensor flies were exposed to cell death-inducing stimuli. The Drosophila ovary is a well-studied cell death model as egg chambers undergo programmed cell death when animals are stressed, a presumed mechanism of matching environmental conditions to progeny production.37,44,45 To induce cell death, biosensor flies were cold-shocked 1 h at -7 °C (freezing the animals), and ovaries were dissected from live animals the next day (Figure 2a). In contrast to untreated biosensor flies, the egg chambers after cold shock were noticeably smaller and exhibited robust red (recent) and green (permanent) biosensor activity, verifying biosensor activation after a cell death stimulus (Figure 2b, merge). Biosensor activity was detected in the nuclei of both germ cells (nurse cells and oocytes) and somatic cells (follicle cells) in egg chambers, but only in the treated flies.35 Morphologies characteristic of apoptosis were also readily observed in biosensor-positive egg chambers including nuclear fragmentation, DNA condensation and degradation (loss of blue stain). Biosensor activity can be detected in the same cells with both nuclear condensation (Hoechst) and active caspases (immunostain), but the most intense staining for active caspases occurred in areas where both nuclear DNA and caspase biosensor signals were diminished or degraded (Figure 2b).46,47 The cell death mechanisms occurring after cold-shock are not well characterized, and other death mechanisms may be at play. However, similar results were obtained following amino acid starvation, which is known to cause apoptosis.35
To determine if CaspaseTracker could detect non-apoptotic caspase activity, the internal organs of healthy 1-day-old flies were meticulously dissected by removing the autofluorescent cuticle and fat. Tissues from a single representative fly were fixed, mounted, stained with Hoechst to mark nuclei and with Phalloidin stain for F-actin to detect muscle cells. In contrast to healthy egg chambers, which lack biosensor activation, the F-actin-stained muscle cells between egg chambers had both red and green biosensor-activity (Figure 3a, inset). However, it is possible that the biosensor also labels other cells. Many other healthy tissues and organs of optimally reared 1-day-old flies exhibited prominent caspase biosensor activity presumably reflecting basal non-apoptotic caspase function (Figure 3a). Biosensor-positive cells appeared morphologically normal and occurred in organized patterns in tissues suggesting that specific subsets of cells activate caspases, for example stripes of biosensor-positive cells wrapping the hindgut (Figure 3a, inset).
The best evidence that CaspaseTracker is a specific reporter of caspases is the lack of signal in flies expressing the control biosensor, which is identical to CaspaseTracker except for a single Asp to Ala amino acid mutation at the obligatory caspase cleavage site, changing DQVD to DQVA (Figure 1b and 3b). Except for an exceptionally rare cell (Figure 3b, green arrow), the only signal observed in the DQVA control biosensor flies are autofluorescent structures, such as mouth parts (Figure 3b) and ingested food in the crop and elsewhere (Figure 3b and 4), which are also observed in non-transgenic parental flies that lack the caspase biosensor.35 The CaspaseTracker-labeled cells occur at regular intervals along the malpighian (kidney) tubules and often exhibit both recent (red) and past (green) caspase activation simultaneously, while many cells in the brain, optic lobes, cardia, crop, midgut and oviduct are either red or green (Figures 4 and 5a).
For evidence that long-lived cells can survive caspase activity, the optic lobe was immunostained for the pan-neuronal marker ELAV (embryonic lethal abnormal vision).48 The nuclei of many neurons were double labeled with nuclear-targeted GFP from CaspaseTracker and ELAV, consistent with long-lived cells using caspases for non-death functions (Figure 5b). Furthermore, if the biosensor is turned off for 10 days (using Gal80ts), biosensor-positive cells persist for the duration in the gut (Figure 6a and b), brain and elsewhere (not shown). Using the caspase biosensor CaspaseTracker, we provide the first overview of basal caspase activity in whole animals.
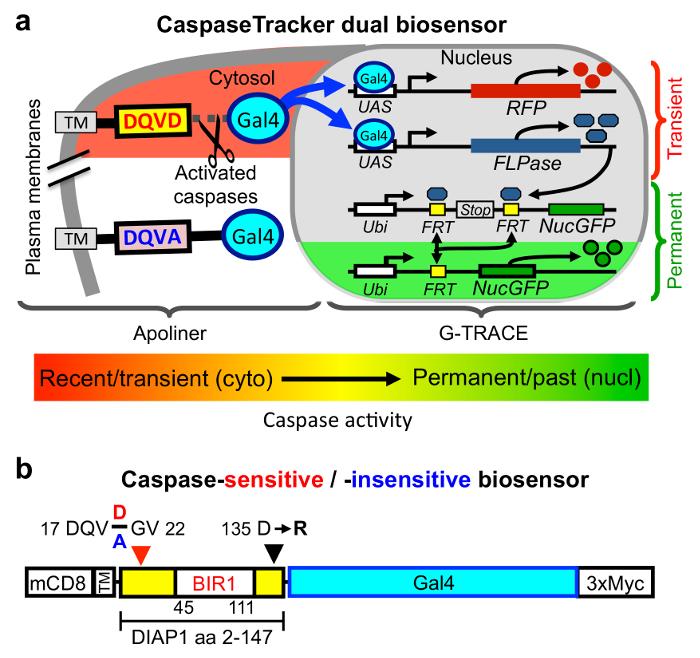
Figure 1: CaspaseTracker biosensor system for detecting non-apoptotic caspase activity. (a) Components of the Drosophila caspase biosensor. (b) The caspase-cleavable portion of CaspaseTracker biosensor system is composed of a plasma membrane anchor (mCD8), a fragment of DIAP1 containing a natural caspase-cleavage site Asp20 fused to the Gal4 transcription factor with a C-terminal 3x-myc tag and expressed via the ubiquitin promoter. Caspase cleavage results in translocation of Gal4 to the nucleus to induce the G-TRACE system. In a control biosensor, the requisite Asp20 residue in the 4-amino acid caspase cleavage site (DQVD) is changed to Ala (DQVA) to abolish caspase cleavage. Transgenic CaspaseTracker flies contain 4 transgenes. Please click here to view a larger version of this figure.
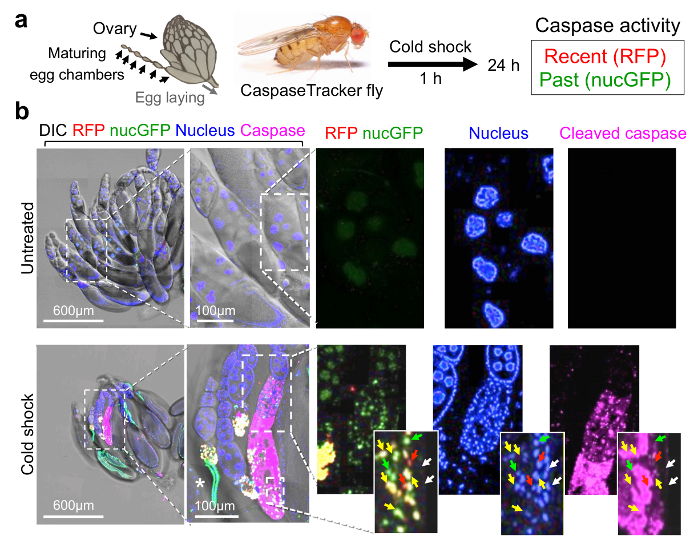
Figure 2: CaspaseTracker is activated by caspases during cell death. (a) Schematic of Drosophila ovary and flow diagram for cold shock-induced cell death. Drosophila ovary drawing (by Polan Santos) and Drosophila image (by Darren Obbard) are used with permission. (b) Egg chambers from the ovary of 6-day female CaspaseTracker flies fed with normal fly food for 6 days (untreated) or 1 day after cold shock (-7 °C, 1 h, followed by 25 °C for 24 h) to induce caspase activation in egg chambers. Enlargements of cold-treated ovary (bottom right frames) show three egg chambers in one developing ovariole chain (centered approximately vertically) with evidence of nuclear condensation (top egg chamber), nuclear fragmentation (middle egg chamber and insets) and degradation (lower egg chamber) based on Hoechst DNA stain (blue). Active caspases (anti-caspase-3, magenta) are detected in the middle and lower egg chambers. RFP and GFP biosensor activity (red, green and yellow arrows) occur in cells with nuclear DNA condensation that often stain diffusely with active caspases (insets). White arrows mark nuclei lacking both biosensor activity and active caspase immunostain. *Autofluorescence signals. These data are reproduced with permission from Tang et al., Sci Rep 2015. Please click here to view a larger version of this figure.
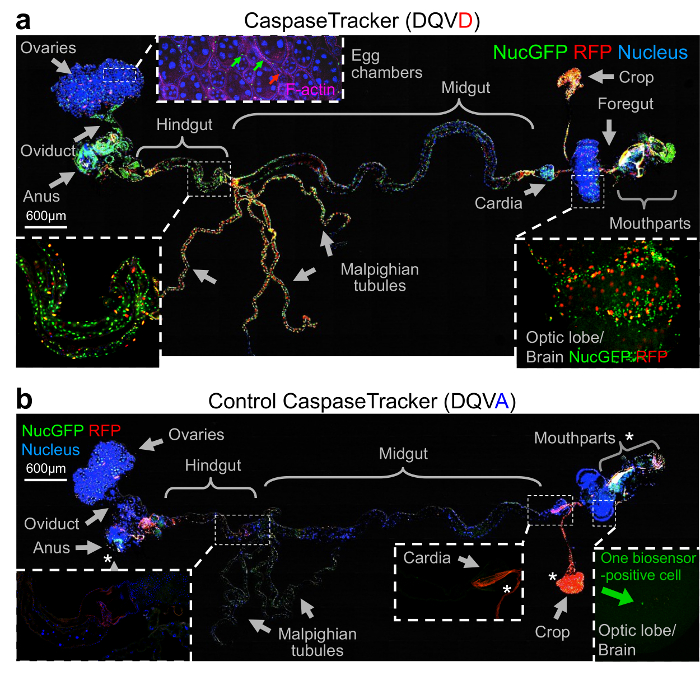
Figure 3: Evidence for widespread physiological caspase activity. (a) Merged confocal image of CaspaseTracker (DQVD) RFP and nucGFP from a single newly eclosed fly raised at 18 °C, dissected and stained with H33342 for nuclei and Phalloidin for F-actin (pink), and imaged with DIC. Phalloidin is shown only in the inset for egg chambers. (b) The same procedure and imaging conditions were followed for control CaspaseTracker (DQVA) flies. *Autofluorescent structures. These data are reproduced with permission from Tang et al., Sci Rep 2015. Please click here to view a larger version of this figure.
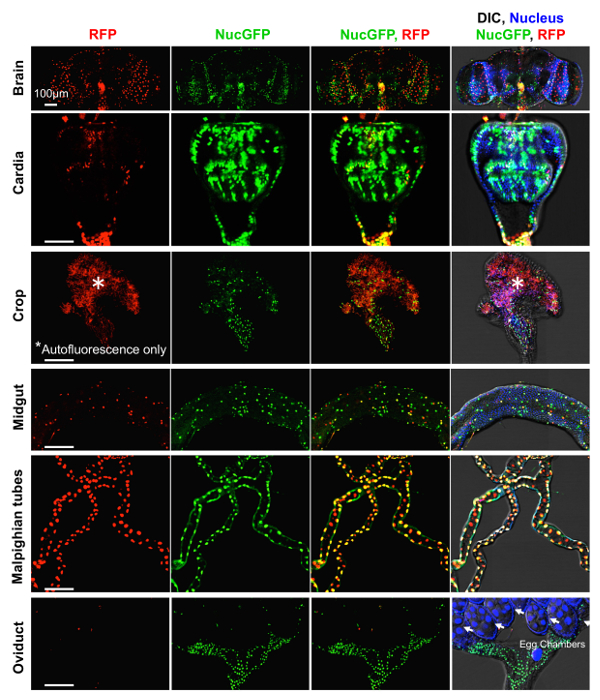
Figure 4: Basal caspase activity in many tissues. Higher magnifications of DIC and fluorescence confocal images of GFP (past) and RFP (recent) CaspaseTracker (DQVD) activity and Hoechst-stained nuclei in the brain, cardia, crop, midgut, Malpighian tubules and oviducts from Figure 3a. *Autofluorescent structures. These data are reproduced with permission from Tang et al., Sci Rep 2015. Please click here to view a larger version of this figure.
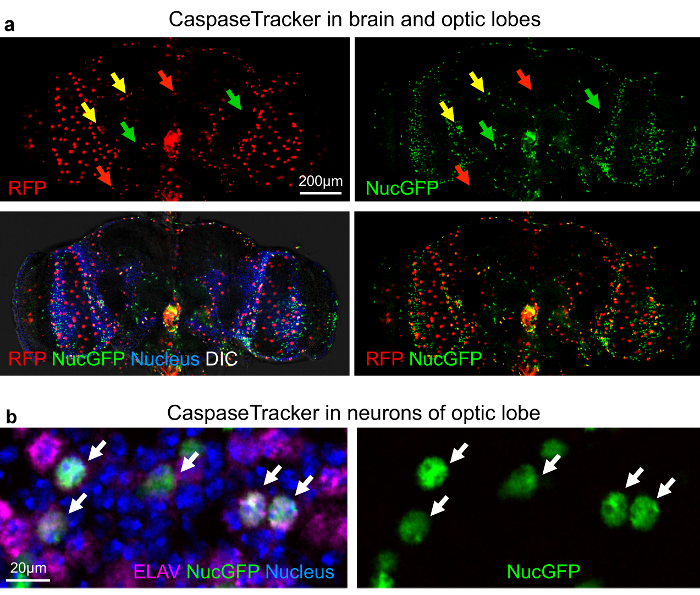
Figure 5: Non-apoptotic caspase activity in neurons of the Drosophila optic lobe. (a) DIC and confocal images with RFP (recent) and GFP (past) CaspaseTracker (DQVD) activity and Hoechst-stained nuclei in the brain. RFP+GFP dual-labeled cells (yellow arrows). (b) NucGFP colocalizes with immunostain for pan neuronal nuclear ELAV protein in the optic lobe of CaspaseTracker (DQVD) fly brain. Neurons with co-localized signals of NucGFP and ELAV are shown (white arrows). Note, RFP-labeled neurons are not present in this particular field. These data are reproduced with permission from Tang et al., Sci Rep 2015. Please click here to view a larger version of this figure.
