To assess β-cell or α-cell replication, a four-color assay protocol is required. First, objects are identified by DAPI staining (Channel 1, 386 nm). Next, β-cells (event 1) are counted: objects that co-express PDX-1+ (channel 2, 650 nm) and peri-nuclear insulin (channel 3, 549 nm). Subsequently, replicating β-cells (event 2) are counted: β-cells (event 1) that co-express Ki-67 (channel 4, 485 nm) (Figure 3). The percentage of replicating β-cells is calculated: (event 2/event 1) x 100. To quantify α-cell replication, total objects are identified by DAPI staining (channel 1) and then α-cells (event 1) are enumerated by the absence of PDX-1 staining (channel 2) and the presence of perinuclear glucagon staining (channel 3). Replicating α-cells (event 2) are the number of α-cells that co-express Ki-67 (channel 4) (Figure 4). The percentage of replicating α-cells is calculated: (event 2/event 1) x 100.
Before initiating the experiment, a compound treatment plan is made (Table 1A). Here, a small scale assay was run using negative control (DMSO-treated) and positive control (dipyridamole-treated [15 µM]) conditions to assess PDX-1, β-cell and α-cell replication in a 384-well format (49 images per well). Experimental data demonstrate dipyridamole to selectively promote PDX-1+-cell and β-cell replication but not α-cell replication. The average number of identified PDX-1-cells per well was 5,541 ± 555 for DMSO-treated and 6,439 ± 363 for dipyridamole-treated conditions (p< 0.01); demonstrating a dipyridamole-dependent increase in PDX-1 cell number (Table 1B, top). The average percent PDX-1+-cell replication (DMSO+PDX-1+Ki-67+/DMSO+PDX-1+) is 3.35 ± 0.50% for DMSO-treated and 10.10 ± 0.98% for dipyridamole-treated conditions (p<0.01) (Table 1B, bottom). Importantly, there are similar PDX-1+-cell replication rates in wells subsequently analyzed for β-cell (rows C-F) and α-cell replication (rows G-J): DMSO= 3.3 ± 0.66 (rows C-F) and 3.4 ± 0.23 (rows G-J) (p= 0.80); dipyridamole= 9.8 ± 0.86 (rows C-F) and 10.4 ± 0.1.1 (rows G-J) (p= 0.43). Hence, these wells, although stained for different cell-lineage markers (insulin and glucagon), responded similarly to drug treatment.
Next the replication rates of β-cells and α-cells were calculated from the raw data (Table 2). The average number of β-cells (event 1) was increased in response to dipyridamole treatment: DMSO 3,930 ± 488 vs. dipyridamole 4,589 ± 218 (p<0.05). Notably, there is a significant reduction in the number of β-cells (3,930 ± 488) compared to PDX-1 cells (5,541 ± 555) (p<0.01). This is a result of excluding PDX-1+-cells that are insulin– from the β-cell count, e.g., δ-cells or β-cell progenitor cells, and the additional stringency of requiring β-cells to be insulin+. Notably, the number of α-cells (event 1) did not increase with dipyridamole treatment (DMSO 1,465 ± 123 vs dipyridamole 1,467 ± 74.7; p= 0.93). Hence, dipyridamole selectively increases β-cell number. The number of replicating β-cells (event 2) is increased in response to dipyridamole treatment (DMSO 131 ± 31 vs. dipyridamole 476.5 ± 39.6; p< 0.01) but the number of replicating α-cells (event 2) is not (DMSO 10.25 ± 3 vs. dipyridamole 9.0±1.6; p=0.5). Finally the percentage of replicating β-cells and α-cells is calculated ((event 2/event 1) x 100). Indeed, dipyridamole increases the percentage of replicating β-cells but not α-cells (summarized in Figure 5). Importantly, the basal β-cell replication rate of a healthy culture varies between experiments (0.5 – 3%) but should be highly consistent across wells within an experiment. Because the basal replication is variable, the compound-induced β-cell replication rate is also variable across experiments; however, the fold-induction of β-cell replication is consistent across experiments and allows compound efficacy to be reliably compared across experiments. Notably, the α-cell replication rate is ~ 5 times lower than the β-cell replication rate and cultures contain ~ 1/3 as many α-cells as β-cells; consequently, the α-cell replication index displays increased variability compared to the β-cell replication index. To limit variability, the images per well is increased from 49 (used here) to a maximum of 81.
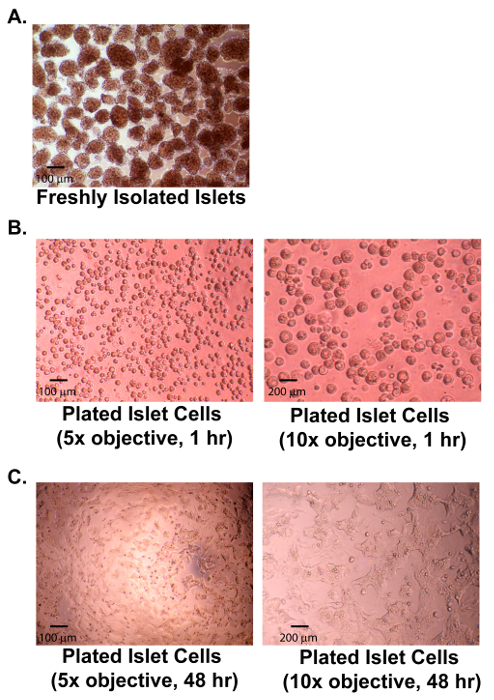
Figure 1: Isolated and Dispersed Rat Islets. (A) A micrograph of healthy isolated rat islets; 100 µm scale bar shown. (B) Micrographs (5X and 10X objectives) of dispersed rat islets 1 hr after plating; 100 µm (left) and 200 µm (right) scale bars shown. (C) Micrographs (5X and 10X objectives) of dispersed rat islets 48 hr after plating; 100 µm (left) and 200 µm (right) scale bars shown. Please click here to view a larger version of this figure.
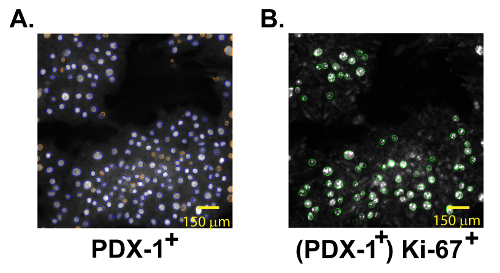
Figure 2: Automated Identification of PDX-1+ and Ki-67+ Islet Cells. (A) Compound-treated dispersed rat islet cells stained for PDX-1. PDX-1+ objects are circled in blue; excluded objects are circled in orange. Scale bar of 150 µm shown. (B) The same field of islet-cells stained for Ki-67. PDX-1+ ki-67+ double-positive cells are circled in green. Scale bar of 150 µm shown. Please click here to view a larger version of this figure.
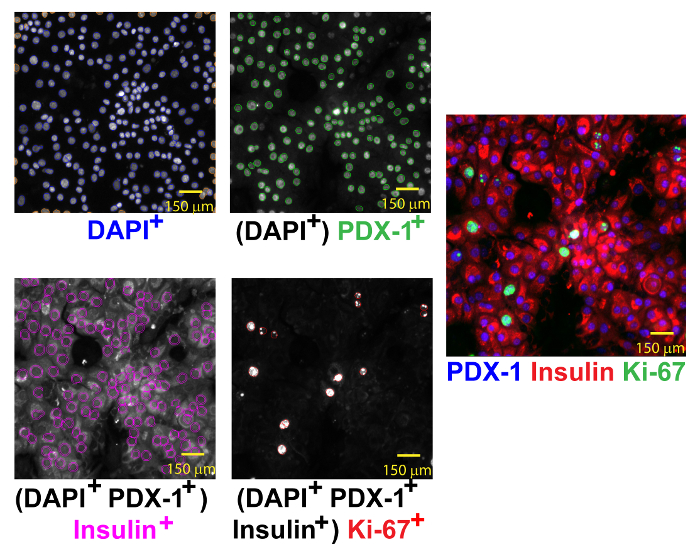
Figure 3: Automated Identification of Replicating β-cells. Replicating β-cells are identified by DAPI (left-upper corner, blue circles), PDX-1 (right-upper corner, green circles), insulin (left-lower corner, magenta circles) and Ki-67 (right-lower corner, red circles) co-expression. The merged image (right) shows several replicating β-cells. Scale bar of 150 µm shown. Please click here to view a larger version of this figure.
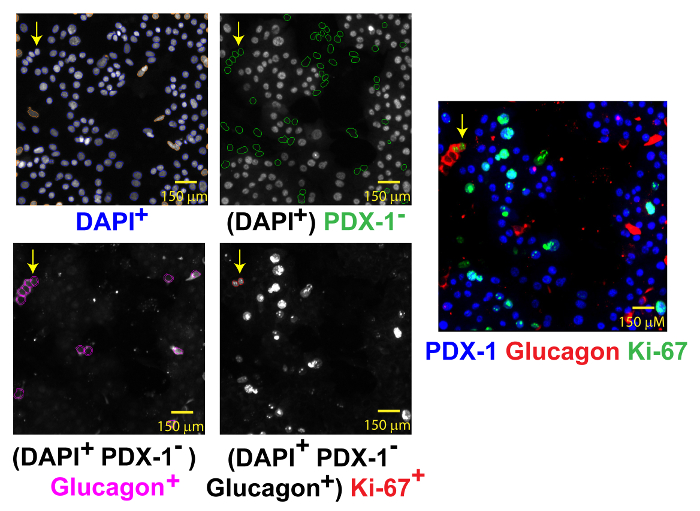
Figure 4: Automated Identification of Replicating α-cells. Replicating α-cells are identified by DAPI (left-upper corner, blue circles), the absence of PDX-1 (right-upper corner, green circles), glucagon (left-lower corner, magenta circles) and Ki-67 (right-lower corner, red circle) co-expression. The merged image (right) shows a rare replicating doublet of α-cells (yellow arrow). Scale bar of 150 µm shown. Please click here to view a larger version of this figure.
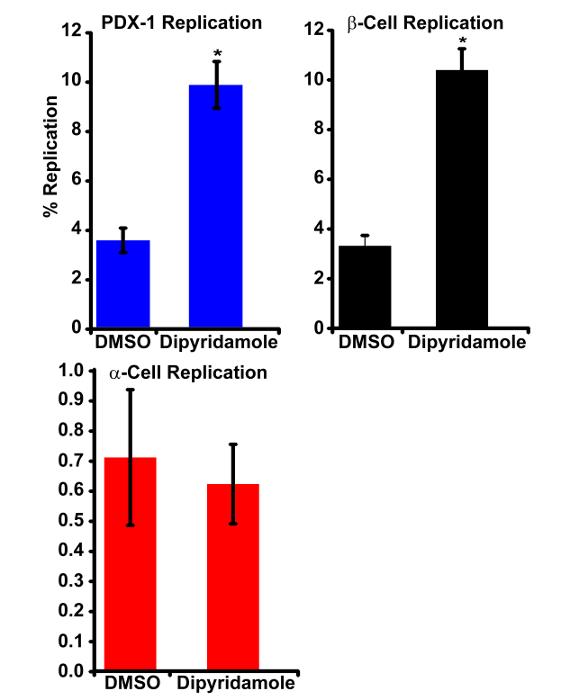
Figure 5: Dipyridamole Induces PDX-1+– and β-cell Replication but Not α-cell Replication. Graphical representation of PDX-1-cell (top left), β-cell (top right) and α-cell (bottom left) replication in DMSO- and dipyridamole-treated wells are shown. Bars represent the mean of independently treated wells (PDX-1 replication, n= 8; β-cell and α-cell replication, n= 4). Error bars indicate the standard deviation (* p< 0.01). Please click here to view a larger version of this figure.
| A. | Treatment: | Staining: | |
| 4 | 5 | ||
| C | DMSO | DIPYRIDAMOLE | Insulin, DAPI, PDX-1, Ki-67 |
| D | DMSO | DIPYRIDAMOLE | Insulin, DAPI, PDX-1, Ki-67 |
| E | DMSO | DIPYRIDAMOLE | Insulin, DAPI, PDX-1, Ki-67 |
| F | DMSO | DIPYRIDAMOLE | Insulin, DAPI, PDX-1, Ki-67 |
| G | DMSO | DIPYRIDAMOLE | Glucagon, DAPI, PDX-1, Ki-67 |
| H | DMSO | DIPYRIDAMOLE | Glucagon, DAPI, PDX-1, Ki-67 |
| I | DMSO | DIPYRIDAMOLE | Glucagon, DAPI, PDX-1, Ki-67 |
| J | DMSO | DIPYRIDAMOLE | Glucagon, DAPI, PDX-1, Ki-67 |
| B. | Number of PDX-1+ Cells | ||
| DMSO | DIPYRIDAMOLE | ||
| 4 | 5 | ||
| C | 5158 | 6249 | |
| D | 6365 | 6795 | |
| E | 4830 | 6974 | |
| F | 5988 | 6408 | |
| G | 5574 | 6532 | |
| H | 5939 | 6411 | |
| I | 5612 | 6387 | |
| J | 4862 | 5759 | |
| Percent of PDX-1+ Ki-67+ Cells | |||
| DMSO | DIPYRIDAMOLE | ||
| 4 | 5 | ||
| C | 3.04 | 9.99 | |
| D | 3.91 | 10.57 | |
| E | 2.51 | 8.58 | |
| F | 3.77 | 10.14 | |
| G | 3.44 | 10.1 | |
| H | 3.06 | 10.69 | |
| I | 3.58 | 9.07 | |
| J | 3.52 | 11.74 | |
Table 1: Representative Experimental Plan and PDX-1+-Cell Replication Data. (A) An outline of the treatment plan is shown. Wells were treated with DMSO (column 4) or dipyridamole (column 5). Staining was performed with DAPI, anti-Pdx-1, anti-insulin (normal lettering) or anti-glucagon (italic lettering) and Ki-67 (B) Total number of PDX-1+ cells (top, DAPI+PDX-1+) per well and the percentage of replicating PDX-1 cells (bottom, DAPI+PDX-1+Ki-67+/DAPI+PDX-1+ x 100) per well are shown.
| Event 1: | |||
| DMSO | DIPYRIDAMOLE | ||
| 4 | 5 | ||
| C | 3557 | 4352 | # DAPI(+) PDX-1(+) Insulin(+) |
| D | 4387 | 4542 | # DAPI(+) PDX-1(+) Insulin(+) |
| E | 3462 | 4879 | # DAPI(+) PDX-1(+) Insulin(+) |
| F | 4315 | 4585 | # DAPI(+) PDX-1(+) Insulin(+) |
| G | 1594 | 1567 | # DAPI(+) PDX-1(+) Glucagon(+) |
| H | 1477 | 1439 | # DAPI(+) PDX-1(+) Glucagon(+) |
| I | 1491 | 1390 | # DAPI(+) PDX-1(+) Glucagon(+) |
| J | 1298 | 1474 | # DAPI(+) PDX-1(+) Glucagon(+) |
| Event 2: | |||
| DMSO | DIPYRIDAMOLE | ||
| 4 | 5 | ||
| C | 113 | 425 | # DAPI(+) PDX-1(+) Insulin(+) Ki-67(+) |
| D | 152 | 511 | # DAPI(+) PDX-1(+) Insulin(+) Ki-67(+) |
| E | 97 | 466 | # DAPI(+) PDX-1(+) Insulin(+) Ki-67(+) |
| F | 163 | 504 | # DAPI(+) PDX-1(+) Insulin(+) Ki-67(+) |
| G | 13 | 9 | # DAPI(+) PDX-1(+) Glucagon(+) Ki-67(+) |
| H | 10 | 9 | # DAPI(+) PDX-1(+) Glucagon(+) Ki-67(+) |
| I | 6 | 11 | # DAPI(+) PDX-1(+) Glucagon(+) Ki-67(+) |
| J | 12 | 7 | # DAPI(+) PDX-1(+) Glucagon(+) Ki-67(+) |
| % Replication: | |||
| DMSO | DIPYRIDAMOLE | ||
| 4 | 5 | ||
| C | 3.18 | 9.77 | |
| D | 3.47 | 11.25 | |
| E | 2.8 | 9.55 | |
| F | 3.78 | 10.99 | |
| G | 0.82 | 0.57 | |
| H | 0.68 | 0.63 | |
| I | 0.4 | 0.79 | |
| J | 0.92 | 0.48 | |
Table 2: Representative β-cell and α-cell Replication Data. The total number of β-cells (Event 1: DAPI+PDX-1+insulin+; top table rows C-F, normal lettering)), α-cells (Event 1: DAPI+PDX-1+glucagon+; top table rows G-H, italic lettering), replicating β-cells (Event 2: DAPI+PDX-1+insulin+Ki-67+; middle table rows C-F, normal lettering) and α-cells (Event 2: DAPI+PDX-1+glucagon+Ki-67+; middle table rows G-H, italic lettering) per well are shown. Additionally, the percentage of replicating β-cells (DAPI+PDX-1+insulin+ Ki-67+/ DAPI+PDX-1+insulin+ x 100; bottom table rows C-F, normal lettering) and α-cells (DAPI+PDX-1+glucagon+Ki-67+/DAPI+PDX-1+glucagon+ x 100; bottom table rows G-J, italic lettering) per well are shown.
