1. Assessing the Effect of Small Molecule Inhibitors on Pellicle and Biofilm Colony Formation
- Prepare a 2x solution of defined biofilm-inducing MSgg medium25 without the calcium chloride and iron(III) chloride hexahydrate. After filter sterilization, add the calcium chloride. The medium is ready to use directly or it can be stored at 4 °C in the dark.
- Prepare the 1x MSgg dilution on the day of the experiment.
- Dilute the 2x MSgg medium to 1x with sterile distilled water (pellicles) or sterile 3% hot (80 °C) agar (biofilms) and add iron(III) chloride hexahydrate to a final concentration of 50 μM (pellicles) or 250 µM (biofilms). Add antibiotics or small molecule inhibitors to the desired concentration and mix well. For example, to obtain a final concentration of 0.5 mM D-leucine in 30 ml to establish pellicles or biofilms, add 196.6 µl of a 76.3 mM (10 mg/ml) D-leucine stock solution.
NOTE: The final 1x MSgg composition is described in Table 1. Compared to the original recipe25, the medium contained 50 µg/ml threonine and the iron concentration to grow biofilm colonies on solid MSgg medium was increased 2.5x to optimize the wrinkled colony morphology.
- Dilute the 2x MSgg medium to 1x with sterile distilled water (pellicles) or sterile 3% hot (80 °C) agar (biofilms) and add iron(III) chloride hexahydrate to a final concentration of 50 μM (pellicles) or 250 µM (biofilms). Add antibiotics or small molecule inhibitors to the desired concentration and mix well. For example, to obtain a final concentration of 0.5 mM D-leucine in 30 ml to establish pellicles or biofilms, add 196.6 µl of a 76.3 mM (10 mg/ml) D-leucine stock solution.
- After solidification of the agar, dry the solid MSgg plates in a biological hood for 30-45 min prior to the inoculation.
- To select specific inhibitors that interfere with the mechanisms of pellicle formation (Figure 1), rule out that the concentrations used affect planktonic and static growth.
- Determine planktonic growth (increase in optical density over time in liquid culture) in a simple growth curve by measuring the optical density at 600 nm every hour until the stationary growth phase.
- To confirm that the measured culture turbidity represents live cell counts, determine the numbers of colony forming units (CFU) of cells in the planktonic growth phase from a shaking culture after several time points.
- To assess the effect of small molecule inhibitors on static pellicle growth, harvest cells at the end of a 3-day incubation at 23 °C from a 24-well cell-culture well, inoculated under the same conditions as described in sections 1.7-1.9 and determine the CFU. For this control, use a pellicle-deficient strain that lacks the operons encoding for the extracellular matrix components (i.e., B. subtilis ΔepsH, ΔtasA).
NOTE: This strain is capable of growing under static conditions, but in contrast to a pellicle-forming wild type, it is deficient in the ability to float to the liquid-air interface, where growth is favored due to increased oxygen levels26. Thus, this extracellular matrix- and pellicle-deficient strain is a recommended reference strain to assess growth under static conditions.
NOTE: For the specific example of the non-canonical D-amino acid D-leucine described below, an effect on planktonic and static growth at concentrations that interfered with pellicle formation was ruled out12,17. The methods to determine planktonic and static growth are described in detail17.
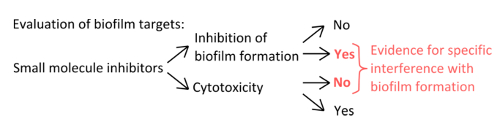
Figure 1. Conceptual overview for the identification of a robust experimental setup to assess the specific inhibition of biofilm formation. Selection criteria for small molecule inhibitors that indicate specific interference with biofilm formation without pronounced effect on planktonic growth. Please click here to view a larger version of this figure.
- Streak out B. subtilis from a -80 °C stock (LB culture of 109 cells/ml frozen in 20% glycerol) to isolate single colonies on a LB-1.5% agar plate with a sterile tip or applicator stick.
- Grow overnight at 30 °C.
- CRITICAL STEP: For a robust pellicle inhibition by the non-canonical D-amino acids such as D-leucine, grow a single colony picked from the LB-1.5% agar plate in 3 ml LB broth at 37 °C for 4 hr in a shaking incubator (shaking speed 200 rpm). Replace the LB broth with biofilm-inducing MSgg medium prior to inoculation by centrifuging 1.5 ml starter culture for 4 min at 6,000 x g, carefully removing the supernatant and re-suspending the pellet in 1.5 ml MSgg medium. The rest of the culture can be discarded.
Important: To ensure the robustness of the system, the optical density at 600 nm (OD600) of the washed starter culture should be between 0.6 and 1. - During the growth of the starter culture, prepare a 12-well cell-culture multidish plate containing 3 ml of MSgg medium without or with a concentration range of the small molecule inhibitor (e.g., 0.3, 0.5, 1 mM D-leucine17). To rule out edge effects, distribute the location of the different concentrations across the multidish plate. Alternatively, use 24-well cell-culture multidish plates containing 1.5 ml of MSgg medium.
- Inoculate the wells of the 12-well cell-culture multidish plate with 3 µl of the washed starter culture (1:1,000 dilution).
NOTE: A lower dilution ratio, i.e., 1:500 can be used. This decreases the development time of the pellicles. - Grow the pellicles at 23 °C under static conditions for three days. Do not move the pellicles during this time, as it can affect the final surface morphology of the pellicle.
- Acquire pictures with a binocular and homogenous exposure of lightning. Alternatively, take a picture of the pellicles with a high resolution camera. To avoid artefacts caused by inconsistent light angles and shadows, take top-down pictures with the camera fixed on a tripod and use a soft and large light source at 45° from both sides.
NOTE: An alternative method to study B. subtilis multicellularity is the biofilm colony assay on solid, biofilm-inducing MSgg medium. Like pellicles, this assay allows the study of spatiotemporal processes. Once the active range of small molecule inhibitors is determined, their effect on biofilm colony formation can be studied. - To grow biofilm colonies, symmetrically spot 1.5 µl of the unwashed pre-culture (Step 1.7) on the dried MSgg 1.5% agar plate with the help of a template — 4 drops per Petri dish of 8.5 cm diameter. Let the drops adsorb to the plate before moving them.
NOTE: The template helps to get an equal distribution of the biofilm colonies within the area where the cells are grown. To prepare the template, draw the total area of growth at original scales, divide it to equal sectors and mark the center. For a round Petri dish of 8.5 cm diameter, this assigns 14 cm2 to one biofilm colony. - Incubate the plates at 30 °C for three days. During this time, biofilm colonies develop and form a three-dimensional, wrinkled structure.
- Take pictures as in step 1.11.
2. Ethanol Resistance Assay
- Grow biofilms as described in steps 1.1-1.7 and 1.12-1.14.
- After 68 hr of growth at 30 °C, cut the biofilm colonies into two equal parts with the help of a razor blade and the template.
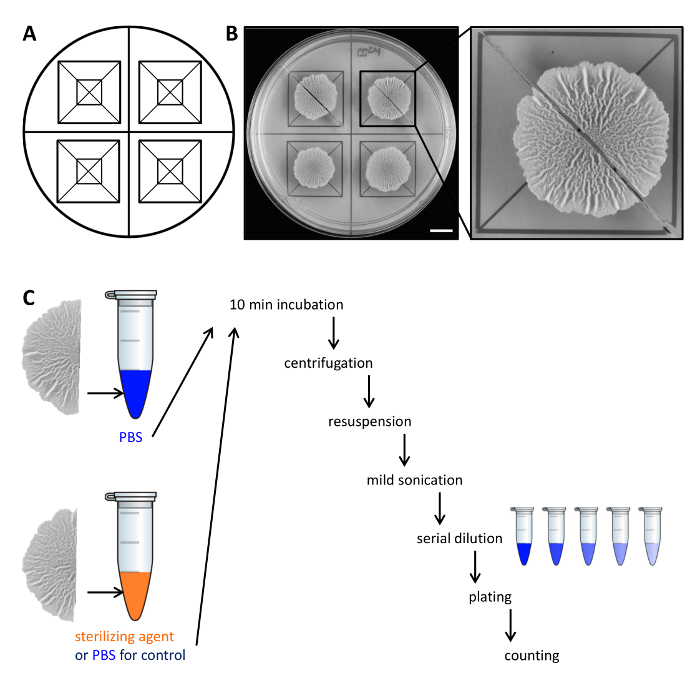
Figure 2. Example experimental design to assess the resistance of biofilm colony cells to sterilizing agents. (A) Template used for the equal distribution of biofilm colonies across a Petri dish and for cutting. (B) Top-down images of untreated wild-type biofilm grown for 68 hr on solid, defined biofilm-inducing MSgg medium at 30 °C. The enlargement shows how a biofilm colony can be cut in two equal halves. (C) The two equal biofilm halves are treated equally (control, PBS) or with either PBS or sterilizing agent and processed as described. Scale bar: 1 cm. Please click here to view a larger version of this figure.
- Carefully lift each half of the biofilm colony from the agar plate with a small spatula and move it to a 1.5 ml microcentrifuge tube containing 500 µl of phosphate-buffered saline (PBS). If necessary, scrape the remaining cells from the plate and transfer them to the microcentrifuge tube as well.
NOTE: The second half of the biofilm colony is treated differentially, depending on whether it is the control or to test the resistance to sterilizing agents. - For the control, incubate the second half of the biofilm colony in 500 µl PBS as in step 2.3. To assess resistance to sterilizing agents, transfer the second half of the biofilm colony to 500 µl 50% (v/v) ethanol.
NOTE: Alternative sterilizing agents such as sodium hypochlorite can be used. For all sterilizing agents used, determine the active concentration and incubation time in a preliminary experiment. - Incubate the biofilm colonies for 10 min on the bench-top at room temperature.
- Centrifuge the biofilm colonies for 5 min at 18,000 x g and carefully remove the supernatant with a pipette. Add 300 µl of PBS.
- Sonicate the cells mildly (amplitude 10%, pulse 5 sec) with the microtip of a sonicator.
NOTE: The sonication energy must be sufficient to separate biofilm aggregates. However, too harsh sonication may lyse the cells. Confirm in advance by light microscopy that the sonication energy used does not lyse the cells and that all aggregates are dissolved. - Add 700 µl of PBS to a final volume of 1 ml. Perform a serial dilution (to 10-7) in PBS and spread 100 µl of 3 dilutions on a LB-1.5% agar plate using sterile glass beads.
NOTE: The optimal dilutions to be plated should be determined in a preliminary experiment, as this depends on the amount of cells in the biofilm colony of interest and the survival rate of the cells in response to the sterilizing agent. - Incubate the plates overnight at 30 °C, count the CFU and determine CFU/ml. From the final CFU/ml of each the half biofilm colonies, calculate the percentage of survivors.
NOTE: When performed and analyzed as described, the two halves of the control biofilm colony and the untreated versus ethanol-treated half of the untreated biofilm colony should yield differences below 10% in viable cell counts, verifying the symmetry or the resistance of the colony, respectively. Alternatively, the results can be represented in total CFU. The cell counts of the control and untreated biofilm colony should remain in the same order of magnitude. In contrast, the cell counts of the ethanol-treated half of a small molecule-treated biofilm colony are expected to drop by a minimum of two orders of magnitude to claim for an increased sensitivity to the sterilizing agent.
3. Biofilm Colony Sample Preparation for Scanning Electron Microscopy
- Grow biofilm colonies as described in steps 1.1-1.7 and 1.12-1.14.
- Prepare a fresh batch of 2% (v/v) glutaraldehyde, 3% (v/v) paraformaldehyde solution in 100 mM sodium cacodylate, 5 mM calcium chloride buffer, pH 7.3. Prepare 5 ml of fixative for every Petri dish of 8.5 cm diameter.
CAUTION: Glutaraldehyde and paraformaldehyde are hazardous. Handle them with safety equipment inside a chemical hood. Discard the solutions and the contaminated materials to the hazardous waste. - Carefully add the fixative to the biofilm colonies, without dispensing directly on top of the biofilms.
NOTE: Due the hydrophobic character of the biofilm colony, the colonies slowly detach from the agar and start to float. - Carefully seal the plates with a strip of Parafilm. Incubate on a rotary shaker for 2 hr at room temperature and subsequently transfer the plates to 4 °C for overnight.
- The next day, carefully remove the liquid with a Pasteur glass pipette connected to a vacuum pump.
- Carefully add 10 ml 100 mM sodium cacodylate, 5 mM calcium chloride buffer to wash the biofilm and incubate for 5 min. Gently remove the liquid with the Pasteur glass pipette from the corner of the plate to avoid damaging the biofilm and add fresh washing solution by gentle pipetting. Repeat this step once.
- For the dehydration of the biofilm colonies, proceed with the following steps: 2x 5 min in ddH2O; 2x 20 min in 30% ethanol; 2x 20 min in 50% ethanol; 2x 20 min in 70% ethanol; 2x 20 min in 96% ethanol; 2x 30 min in 100% ethanol.
- Add 15 ml of liquid for every Petri dish of 8.5 cm diameter in each step and remove the liquid carefully after each incubation.
- Use one of two different methods for drying the samples from ethanol.
- For air-drying from ethanol:
- Cut a cellulose filter paper (diameter of 9 cm) in quarters. Briefly submerge one quarter in 100% ethanol, and then carefully transfer one floating biofilm colony onto it. Put the wet filter paper in a Petri dish lined with a filter paper. Cover the Petri dish and let the biofilm colonies dry overnight in a chemical hood.
- For critical point (CP)-drying using carbon dioxide (CO2) as the transition fluid:
- Fill 75% of the critical point drying machine chamber with 100% ethanol. Transfer the samples into a holder, each sample into its own chamber. If necessary, cut the biofilm with scissors into smaller pieces. Leave the samples submerged in ethanol during all the handling. Then, transfer the holder into the chamber and close the chamber tightly.
- Cool the chamber to 7 °C and start stirring. Fill the chamber completely with liquid CO2. During a 7 min incubation time, let the ethanol mix with the CO2. Then, discharge 25% of the solution.
NOTE: Do not empty the chamber below the level of the sample. - Repeat step 3.8.2.2 four times.
- Repeat step 3.8.2.2 five times with an incubation time of 5 min only. Finally, the ethanol should be completely replaced by CO2.
- During the last round, empty only 5% of the chamber. Turn off the stirring and the cooling. Start heating the chamber to 42 °C. At a temperature of 31.1 °C and a pressure of 73.9 bar, the liquid CO2 reaches its critical point, the state where the gaseous phase has the same density as the liquid phase of the solvent27. Once the temperature reaches 42 °C, incubate for 10 min. At 42 °C, the CO2 in the chamber exists as supercritical gas.
NOTE: Constantly check the pressure of the chamber. The pressure should not exceed 120 bar at 42 °C. - Start to slowly release the gas with continuous heating. This keeps the samples in the CO2-gas phase and prevents the deformation of the sample morphology through the liquid surface tension. Set the flowmeter to 5 L/hr by fine tuning the metering valve controlling the flowmeter. Wait until the all the pressure in the chamber is released. Now open the chamber and remove the samples carefully from the holder.
- For air-drying from ethanol:
- Coat an electron microscopy stub with carbon tape. With the help of tweezers, carefully transfer the biofilm colonies onto the stub. Connect each colony to the stub by adding a thin bridge from carbon tape, which is crucial for charge elimination under the electron beam. At this stage, handle the biofilm colonies with care as they are very fragile. Store the samples in a desiccator for at least 24 hr or until examination.
- The day of the examination with the scanning electron microscope, sputter-coat the biofilm colonies for 2 min in a 60° angle in a gold-palladium sputter-coater. Repeat this step twice and rotate the samples by 120° in between. At the end, sputter-coat the samples once for 3 min from the top. The 20 nm thin layer of gold-palladium improves the conductivity and enhances the contrast of the sample for the imaging in the SEM.
- Store the samples in a desiccator to avoid the rehydration of the sample28 until imaging with a scanning electron microscope29,30.
The pellicle assay is one method to study the highly regulated and dynamic processes of B. subtilis multicellularity. Besides this, the pellicle assay is suited to test a range of either pre-starter conditions or small molecule concentrations in a single cell-culture multidish plate in one experiment. However, B. subtilis pellicle formation is sensitive to the pre-culture conditions (e.g., growth medium of the pre-culture and its growth phase), the inoculation ratio and the removal of the pre-culture medium. We therefore first screened for an experimental setup that allowed us to reproduce pellicle inhibition by D-leucine. Our results show that reproducible pellicle inhibition by D-leucine can be obtained if the cells are pre-cultured in undefined, rich LB medium to a mid-logarithmic growth phase (single colony in 3 ml LB at 37 °C for 4 hr with shaking), spun down and re-suspended in defined MSgg medium prior to a 1:1,000 inoculation (Figure 3A). When the cells were grown in MSgg medium to the mid-logarithmic growth phase (single colony in 1 ml LB for 2 hr, re-diluted 1:100 to 3 ml MSgg, and grown for 5 hr at 37 °C with shaking at 200 rpm), removal of the pre-culture medium did not have an effect on D-leucine activity. Cells grown to the stationary growth phase (single colony in 1 ml LB for 2 hr, re-diluted 1:100 to 3 ml MSgg, and grown for up to 20 hr in a roller at room temperature) and washed in MSgg medium prior to inoculation were less sensitive. However, this increase in resistance towards the pellicle inhibition effect of D-leucine could be due to the higher cell density of the pre-culture, increasing the inoculation ratio. The inoculation ratio of the pre-culture to the static growth in MSgg medium, i.e., 1:500 or 1:1,000, influenced the activity range of D-leucine and the time of pellicle development (Figure 3B). Pellicle inhibition by D-leucine occurred at different growth temperatures (23 °C and 30 °C, Figure 3C). Importantly, while the sensitivity of cells to D-leucine depends on the pre-culture conditions, the reproducibility of the phenomena at different temperatures demonstrates that pellicle inhibition by the small molecule D-leucine has robust features.
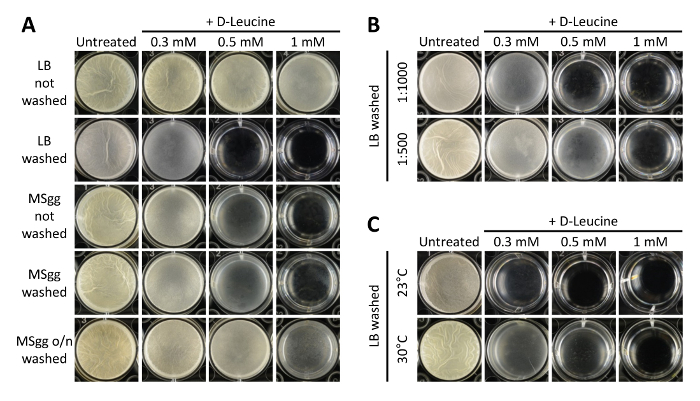
Figure 3. Example results that show the effects of various pre-culture conditions on pellicle inhibition by D-leucine. Several parameters have to be assessed to obtain a robust experimental setup, including: (A) removal of contaminants from the pre-culture growth medium (not washed versus washed), growth medium of the pre-culture (rich, undefined LB medium or defined biofilm-inducing MSgg medium), and growth state of the pre-culture (logarithmic versus stationary growth phase), (B) inoculation ratio (1:1,000 versus 1:500) of the pre-culture into the final pellicle growth medium, and (C) growth temperature (23 °C versus 30 °C). B. subtilis NCIB 3610 cells were grown in the medium indicated (LB or MSgg) for 4 hr at 37 °C or at room temperature over night (o/n) with shaking, and pre-culture medium was replaced by MSgg if indicated (washed or not-washed). Finally, the starter culture was inoculated 1:1,000 if not stated otherwise and pellicles were grown under static conditions at 23 °C for three days. Top-down photographs were acquired with a camera. Well diameter is 22 mm. Please click here to view a larger version of this figure.
After the identification of pre-culture conditions and an experimental setup that allowed for a reproducible activity range of D-leucine on pellicle formation (pre-culture grown in LB to a mid-logarithmic growth phase, washed in MSgg, diluted 1:1,000 and grown at 23 °C for three days), the effect was tested for its robustness. For that purpose, the activity of D-leucine on pellicle formation was assessed in various modified MSgg media (Figure 4). The activity range of D-leucine in pellicle inhibition was consistent in the five media tested, and shows that the effect of D-leucine on pellicle formation is robust. A similar trend was observed with D-Tyrosine, where pellicle inhibition in the concentration range up to 2 µM was consistent in several modified MSgg media (lower iron concentration and replacement of the nitrogen source with ammonium chloride or the carbon source with glucose, data not shown).

Figure 4. Sensitivity to D-leucine occurs in various growth media. Shown are top-down photographs of B. subtilis NCIB 3610 pellicles, grown under static conditions at 23 °C in defined, biofilm-inducing MSgg medium or in different modified MSgg media (carbon source 0.5% glucose; nitrogen source 0.5% (NH4)2SO4; high iron 250 µM FeCl3; low glycerol 0.125% glycerol) for three days, except for the alternative nitrogen source medium (where the pellicles were grown for five days) either without or with D-leucine at concentrations indicated. Starter cultures were grown for 4 hr at 37 °C with shaking in undefined, rich LB medium. Prior to inoculation, cells were washed by centrifugation and resuspended in the corresponding pellicle growth medium. The starter culture was diluted 1:1,000, the well diameter is 22 mm. Please click here to view a larger version of this figure.
An alternative method to study B. subtilis multicellularity is the biofilm colony assay on solid MSgg medium. Like pellicles, this assay allows the study of spatiotemporal processes. Once the active range of small molecule inhibitors is determined in the pellicle system, their effect on biofilm colony formation can be studied. In this study, biofilm colonies were used to assess the effect of D-leucine on the resistance of biofilm colonies to sterilizing agents, because biofilm colonies are less fragile and on the solid background, they are easier to manipulate. When biofilm colonies treated with D-leucine are exposed to 50% ethanol for 10 min, the percentage of surviving cells drops dramatically compared to the untreated fraction (Figure 5). This method can be developed further to assess the effects of other small molecule inhibitors and their effect on the resistance to bactericidal agents (sterilizing agents or antibiotics).
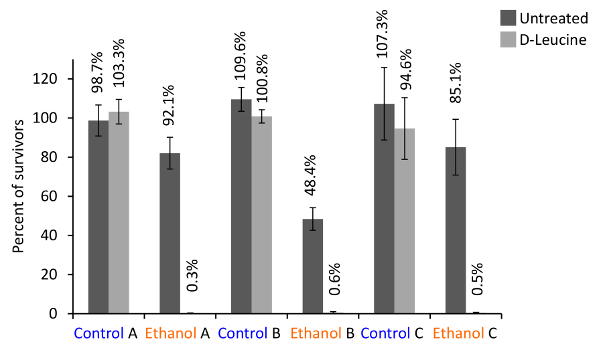
Figure 5. Example results of untreated or treated biofilm colony resistance against a sterilizing agent. Biofilm colonies of B. subtilis NCIB 3610 were grown on solid, defined medium for 68 hr at 30 °C, with or without 0.5 mM D-leucine. After cutting the colonies in two equal halves, one half was treated with PBS (internal control) and the other half with 50% ethanol or PBS (control). After mild sonication, serial dilution, plating and counting of the colony forming units, the percentage of survivors compared to the internal control was calculated. Shown are the results of three independent experiments with the averages of two (A) or three (B, C) replicates and their standard deviations. Please click here to view a larger version of this figure.
Scanning electron microscopy (SEM) is a powerful tool that can be used to focus on the spatial architecture of biofilm colony wrinkles, the abundance and localization of the EPS and single cell morphology31. SEM imaging requires samples that can be imaged under high vacuum. To investigate samples under high vacuum conditions, all bulk water molecules have to be removed, and therefore, SEM imaging requires the dehydration of the sample prior to imaging. Alternatively, samples can be imaged under low vacuum conditions in the wet mode by environmental scanning electron microscopy (ESEM), where the hydrated sample is gently dried in the microscope chamber. To evaluate the effects of small molecule inhibitors on biofilm colony architecture, the EPS organization and cell morphology, three different types of sample dehydration were compared: drying in the microscope under wet mode conditions (ESEM), air-dried from a solvent or critical point (CP)-dried using carbon dioxide as the transition fluid. In untreated and D-leucine-treated B. subtilis biofilm colonies different hydration stages according to the method used could be observed. Imaging under low vacuum conditions in the wet mode by ESEM preserved the very natural state of the biofilm colony. However, information about single cell morphology and EPS abundance and architecture was scarce, as the EPS were still fully hydrated and the cells were embedded in the EPS (data not shown). In terms of dehydration, CP-drying was the most stringent procedure. It removed most structurally bound and bulk water molecules from the tissue, accounting for a volume loss of 30-70%, depending on the tissue32. In contrast, air-drying preserved the bound water molecules. An embryonic tissue for example lost 18% of its volume after air-drying from the volatile solvent ethanol and 59% after CP-drying32. As the EPS bind water and are very hydrated, we recommend considering the complete loss of water in the CP-dried method. When the samples of this study were dehydrated in ethanol and air-dried, the three-dimensional structure of the biofilm colony was preserved, single cells appeared, and the EPS were flattened, but they still connected and covered the single cells (Figure 6A). Biofilm colonies that were dehydrated with ethanol and CP-dried also preserved their three-dimensional structure, and the EPS appeared like a spider web (Figure 6C). In the air-dried and CP-dried samples, the effects of the small molecule inhibitor D-leucine on the biofilm colony architecture, the single cell morphology and EPS structure were apparent (Figure 6B and D). The biofilm colonies grown in presence of D-leucine were smaller and the wrinkles were less pronounced. The D-leucine-treated cells were elongated, a phenotype that is observed in cells treated with cell wall-targeting molecules33. Additionally, the cells were clearly less covered with the EPS and not tightly connected to their neighboring cells via the EPS as seen in the untreated biofilm colonies. The presented results show that both methods of biofilm colony dehydration, air-dried from ethanol or CP-dried using carbon dioxide as the transition fluid give valuable insights on the effect of small molecules on the single cell morphology, as well as on the EPS abundance and architecture.
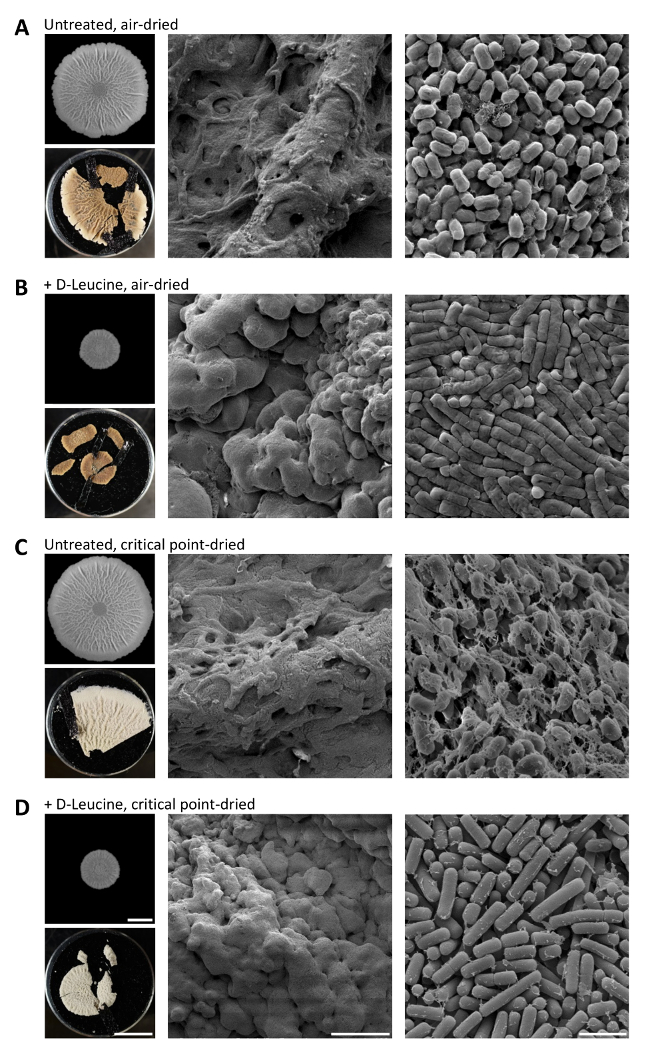
Figure 6. Small molecules can change the large and small scale of the biofilm colony architecture. Biofilm colonies of B. subtilis NCIB 3610 grown (A and D) in absence or (B and D) in presence of 0.5 mM D-leucine for 72 hr at 30 °C on solid MSgg medium were fixated as described in the protocol and dried from ethanol as following: (A and B) air-dried and (C and D) CP-dried using carbon dioxide as the transition fluid. Shown are top down binocular images of the biofilm colonies (left, upper panels), top down photographs of the dried biofilm colonies mounted on electron microscopy stubs prior to gold-palladium-coating (left, lower panels), low- and high-magnification images acquired by SEM to show the 3D architecture of the biofilm colony wrinkles or the single cell morphology/EPS organization. Scale bars from left to right: 5 mm, 100 µm, 2 µm. Please click here to view a larger version of this figure.
| 1.925 mM | potassium phosphate monobasic |
| 3.075 mM | potassium phosphate dibasic |
| 100 mM | 3-(N-morpholino)propanesulfonic acid, pH 7.1 |
| 2 mM | magnesium chloride hexahydrate |
| 700 μM | calcium chloride anhydrous |
| 50 μMa | iron(III) chloride hexahydrate |
| 125 μMb | |
| 1 μM | zinc chloride anhydrous |
| 2 μM | thiamine hydrochloride |
| 50 μg/ml | tryptophan |
| 50 μg/ml | phenylalanine |
| 50 μg/ml | threonine |
| 0.5% (v/v) | glycerol anhydrous |
| 0.5% (w/v) | L-glutamic acid monosodium salts hydrate |
| afor pellicle assay; bfor biofilm assay | |
Table 1. Final 1x MSgg composition used in this study.
