الخلايا تحتوي على العديد من الجزيئات الكبيرة التي تتفاعل باستمرار مع بعضها البعض. هذا الارتباط يثير المجمعات التي تشارك في مسارات الخلوية المسؤولة عن عملها في نقل الإشارة، وتنظيم التعبير الجيني وهجرة الخلايا وغيرها. تشمل جميع التفاعلات البروتين البروتين التي تحدث في خلية شبكة تعرف باسم interactome. في خميرة الخباز وقد تبين أن أكثر من 70٪ من البروتينات لدينا شركاء التفاعل 1. فهم interactome من خلية وتوفير وظائف لهم المعلومات ذات الصلة على مدى تعقيد وتنوع الكائنات الحية. وقد وصفت عدة منهجيات لتحديد وتوصيف التفاعلات البروتين البروتين. تختلف عالية من خلال طرق وضع مثل الخميرة اثنين الهجين 2، البروتين جزء المقايسات تكامل 3، تقارب تنقية 4 بالإضافة إلى قياس الطيف الكتلي وmicroarra البروتينتستخدم يس لتحديد التفاعل 5،6. وبمجرد تحديدها، فمن الضروري التحقق من صحة ذلك، وهذا قد تختلف على أساس كل حالة على حدة. عادة، هذه التجارب تنطوي على تعطيل التفاعل نفسه في مستوى أفراد الزوج التفاعل، على سبيل المثال، عن طريق حذف الجينات أو overexpression من واحد من البروتينات، وبعد ذلك تبحث عن التغيرات في خصائص أو وظيفة عضو آخر في المستوى الخلوي. وفي وقت لاحق، وتستخدم التقنيات الفيزيائية الحيوية 7 لوصف التفاعل البروتين البروتين على المستوى الجزيئي. تحقيقا لهذه الغاية، ويتم تحديد بنية البروتين المجمعات التي كتبها البلورات بالأشعة السينية، الرنين المغناطيسي النووي والفحص المجهري البرد الإلكترون في حين تستخدم الكالوري ومضان الطيفي لكميا وميكانيكيا وصفها.
في هذا العمل، وقد استخدم مضان تباين كأسلوب لوصف التفاعل بين GTPase EFL1 وSBDS البروتين. تشارك هذه البروتينات في تركيب الريبوسومات من خلال تشجيع إطلاق سراح حقيقية النواة عامل بدء 6 من سطح 60S الوحيدات الريباسي 8. وتحور البروتين SBDS في المرض المعروف باسم Shwachman الماس متلازمة 9 و بمثابة عامل الصرف جوانين النوكليوتيدات لEFL1 انخفاض تقارب من أجل غوانوزين ثنائي فسفات 10،11. الطفرات المرض في SBDS تلغي التفاعل مع EFL1 وبالتالي تمنع تفعيله.
يستخدم مضان تباين شيوعا في التطبيقات البيولوجية لدراسة بروتين الببتيد أو تفاعلات حمض البروتين النووي. لأنه يقوم على مبدأ أن fluorophore منتهى السعادة لعودته وكانت النتائج الضوء المستقطب في انبعاث الاستقطاب جزئيا. ويعرف تباين مضان من المعادلة 1:
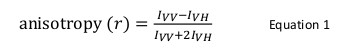
حيث أنا VV وأنا VH هيشدة مضان من عموديا (VV) وأفقيا (VH) الاستقطاب الانبعاثات عند العينة هو متحمس مع الاستقطاب عموديا ضوء 12. مضان تباين حساسة للعوامل التي تؤثر على معدل انتشار التناوب من fluorophore ويعتمد على درجة الحرارة، واللزوجة من الحل وحجم الجزيئي واضح من fluorophore الآن. حجم واضح من البروتين تحتوي على زيادات fluorophore عندما يتفاعل مع بروتين آخر وهذا التغيير ومن ثم يمكن تقييمها على أنها تغير في الخواص المتباينة. وبشكل أكثر تحديدا، فإن fluorophore التي تدور ببطء في حل قريب لعمر الفلورسنت التي يكون لها قيمة أنا VV الكبيرة والصغيرة القيمة أنا VH وبالتالي ستعرض على تباين واسع نسبيا. لfluorophores أن تعثر بسرعة بالنسبة لحياتهم الفلورسنت، وأنا VV وأنا VH تكون مشابهة وسوف قيمة تباين على أن تكون صغيرة 12 </sup > (الشكل 1). وبالإضافة إلى ذلك، إشارة تباين جيدة لقياس الضوضاء، فمن الضروري أن يكون هناك fluorophore مع عمر مضان مماثل للوقت ارتباط التناوب للجزيء من الفائدة. خلاف ذلك، فإنه ليس من الممكن أن يسجل بدقة الفرق في تباين بين البروتين مجانا وأنه في المجمع. على سبيل المثال، وتباين في تحقيق الفلورسنت مع عمر قريب من 4 NSEC مثل فلوريسئين أو رودامين تعلق على انخفاض الوزن الجزيئي مجمع 100 دا هو 0.05. ملزمة لجزيء من 160 كيلو دالتون وزيادة قيمة تباين ل0.29. الفرق التي يمكن قياسها بدقة. في المقابل، فإن نفس التحقيق الفلورسنت تشارك في رد فعل ملزم الذين الزيادة في حجم الجزيئي يختلف من 65 إلى 1000 كيلو دالتون يؤدي إلا إلى تغيير تباين من ،28-،3، التي هي صغيرة جدا بحيث لا يمكن قياسها بدقة. في هذا السيناريو، فإن التحقيق مع عمر من 400 NSEC أن يكون أكثر مناسبة 12.
<p الطبقة = "jove_content">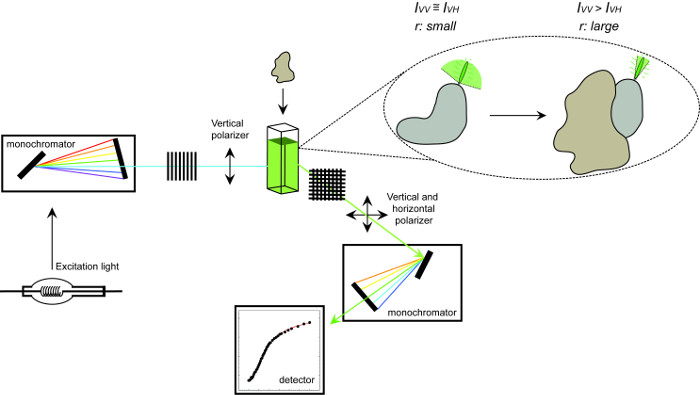
الشكل 1. تمثيل تخطيطي من المعدات المستخدمة لقياس تباين مضان والإجراء. تمثيل تخطيطي من المعدات المستخدمة لإجراء البروتين البروتين التفاعل التجربة قياس تباين مضان. Fluorophores أن تعثر سريع عرض تباين الصغيرة التي يزيد عليها ملزمة لشريك التفاعل. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
تتطلب التطبيقات مضان وجود fluorophore في أي من الجزيئات التي شملتها الدراسة. لدراسة تفاعلات البروتين البروتين هناك ثلاثة أنواع من fluorophores: 1) بقايا التربتوفان موجودة في البروتينات، 2) fluorophores تعلق كيميائيا و 3) شركاء الانصهار فلوري مثل البروتين الفلوري الأخضر (GFP) وderiva لهاالأقارب. معظم البروتينات لها بقايا التربتوفان على هيكلها، وبالتالي فإن أسهل طريقة لقياس التفاعل هو عن طريق رصد التغيرات في أطياف مضان المقابلة أو عن طريق رصد التغيرات في كثافة مضان من بقايا التربتوفان. ومع ذلك، قد تكون بقايا التربتوفان الحالي في كل من البروتينات تعقيد التحليل. من ناحية أخرى، لfluorophore لتغيير خصائص فلوري لها بسبب التفاعل فإنه يحتاج إلى أن يكون موجودا على أو بالقرب من موقع ملزمة وأنها يمكن أن تتداخل مع التفاعل نفسها. هذا يحتاج إلى عناية خاصة عند استخدام fluorophores ضخمة مثل GFP. إذا كان أي من هذه fluorophores يمكن استخدامها لدراسات ملزمة من الضروري، بعد ذلك، إلى إدخال fluorophores خارجي إلى واحد من البروتينات ذات الصلة. وجود العديد من fluorophores تصنيعه كيميائيا ويمكن أن تعلق تساهمي للبروتينات من خلال مجموعاتهم التفاعلية مثل المجموعات أمين (سلسلة من الجانب lysines أو N-محطة) ومجموعة ثيول في السيستين. Fالمشتقات luorophore مع ثيوسيانات وsuccinimidyl استرات تتفاعل مع مجموعة أميد في حين iodoacetamide وmaleimide مجموعات ثيول-رد الفعل (13). الأصباغ الأكثر شيوعا في التطبيقات مضان هي مشتقات من فلوريسئين والأصباغ الخضراء رودامين، الكومارين، fluorophores BODIPY والأصباغ اليكسا فلور. ويمكن الاطلاع على قائمة مفصلة من fluorophores المتاحة تجاريا واستخدامها في المراجع 14،15. لوضع العلامات ناجحة، يجب أن تتعرض لمجموعة رد الفعل على سطح البروتين، ولكن نظرا لوجود عدد كبير من المجموعات الوظيفية على رد الفعل الحالية عادة في البروتينية من الصعب جدا الحصول على تعديل للموقع المحدد. البروتين من الاهتمام في هذه الدراسة، SBDS، ويحتوي على 5 cysteines مجانية و 33 lysines التي قد تؤدي إلى العديد من العلامات الموقع. وضع العلامات غير موحدة قد يؤثر على ملزمة وسيعقد تحليل البيانات والجزيئات fluorophore المختلفة قد تثير مختلفة إشارات كثافة الفلورسنت على ملزمة. لاوفهrcome هذه المشكلة، استخدمنا fluorophore فلاش، 4، 5'مكرر (1،3،2 dithioarsolan-2-YL) فلوريسئين لتسمية المباشر الموقع البروتين SBDS. هذا هو صبغ أرسينوكسيد مع قابلية عالية لمدة أربعة cysteines متباعدة في عزر تعرف باسم فلاش العلامة تتكون من CCXXCC تسلسل حيث X هو أي حمض أميني غير السيستين 16،17. يضاف هذا الحافز tetracysteine إلى N- أو C-محطة من البروتين عن طريق الهندسة الوراثية جنبا إلى جنب مع رابط المناسبة لمنع اختلال أضعاف العام للبروتين. وقد تم تصميم هذا الزوج تتكون من فلاش صبغ وفلاش العلامة أصلا للبروتينات تسمية مواقع محددة في الخلايا الحية 17 ولكن يمكن أيضا أن تستخدم لتسمية تنقية البروتينات في المختبر كما يتجلى ذلك هنا. بالإضافة إلى ذلك، كما تم وضع استراتيجيات الأنزيمية لتمكين functionalization في مواقع محددة من البروتينات 18.
في هذه المخطوطة وصفنا فائدة مضان تباين لأداة سا لدراسة تفاعلات البروتين البروتين. ملزمة يمكن تقييمها عن طريق التفتيش بسيط من شكل منحنى ملزم بينما يمكن الحصول على معلومات كمية من نوبة من البيانات التجريبية.
