प्रकोष्ठों Biomacromolecules कि लगातार एक दूसरे के साथ बातचीत के एक भीड़ में होते हैं। इस संस्था परिसरों कि संकेत पारगमन, जीन अभिव्यक्ति और अन्य लोगों के बीच सेल प्रवास के नियमन में उनके कामकाज के लिए जिम्मेदार सेलुलर रास्ते में भाग लेने के लिए जन्म देता है। सभी प्रोटीन, प्रोटीन बातचीत है कि एक सेल में घटित एक नेटवर्क interactome के रूप में जाना शामिल हैं। Saccharomyces cerevisiae में इसकी प्रोटीन की 70% से अधिक बातचीत भागीदारों 1 है दिखाया गया है। एक सेल के interactome को समझना और उनके कार्यों जटिलता और रहने वाले जीवों की विविधता पर प्रासंगिक जानकारी प्रदान करते हैं। कई तरीके की पहचान करने और प्रोटीन, प्रोटीन बातचीत को चिह्नित करने वर्णित किया गया है। ऐसे खमीर दो संकर 2, प्रोटीन-टुकड़ा पूरक assays 3, आत्मीयता शुद्धि 4 मास स्पेक्ट्रोमेट्री और प्रोटीन microarra के लिए युग्मित के रूप में डाल तरीकों के माध्यम से विभिन्न उच्चवाईएस एक बातचीत 5,6 की पहचान करने के लिए इस्तेमाल कर रहे हैं। एक बार की पहचान, यह यह मान्य करने के लिए आवश्यक है और यह एक मामला-दर-मामला आधार पर भिन्न हो सकते हैं। आमतौर पर, इन प्रयोगों बातचीत जोड़ी, जैसे के व्यक्तिगत सदस्यों के स्तर पर बातचीत से ही खलल न डालें, जीन विलोपन या प्रोटीन में से एक की overexpression से शामिल है, और फिर कम से गुणों में परिवर्तन या अन्य सदस्य के समारोह के लिए तलाश सेलुलर स्तर। बाद में, biophysical तकनीक 7 आण्विक स्तर पर प्रोटीन, प्रोटीन बातचीत को चिह्नित करने के लिए इस्तेमाल कर रहे हैं। यह अंत करने के लिए, प्रोटीन परिसरों की संरचना एक्स-रे क्रिस्टलोग्राफी, परमाणु चुंबकीय अनुनाद और क्रायो इलेक्ट्रॉन माइक्रोस्कोपी द्वारा निर्धारित कर रहे हैं, जबकि उष्मामिति और प्रतिदीप्ति स्पेक्ट्रोस्कोपी मात्रात्मक और mechanistically उन्हें का वर्णन करने के लिए इस्तेमाल कर रहे हैं।
इस काम में, प्रतिदीप्ति anisotropy GTPase EFL1 और SBD के बीच बातचीत को चिह्नित करने के लिए एक तकनीक के रूप में इस्तेमाल किया गया थाएस प्रोटीन। ये प्रोटीन 60 ribosomal सबयूनिट 8 की सतह से यूकेरियोटिक दीक्षा कारक है 6 की रिहाई को बढ़ावा देने से राइबोसोम के संश्लेषण में भाग लेते हैं। SBDS प्रोटीन एक बीमारी EFL1 guanosine diphosphate 10,11 के लिए अपने संबंध को कम करने के लिए एक गुआनिन न्यूक्लियोटाइड विनिमय कारक के रूप में Shwachman-डायमंड सिंड्रोम 9 और कृत्यों के रूप में जाना जाता है में उत्परिवर्तित है। SBDS में रोग म्यूटेशन EFL1 के साथ बातचीत को खत्म करने और इस प्रकार अपने सक्रियण को रोकने के।
प्रतिदीप्ति anisotropy सामान्यतः प्रोटीन पेप्टाइड या प्रोटीन-न्यूक्लिक एसिड बातचीत का अध्ययन करने के लिए जैविक अनुप्रयोगों में प्रयोग किया जाता है। यह सिद्धांत है कि एक fluorophore एक आंशिक रूप से ध्रुवीकृत उत्सर्जन में ध्रुवीकृत प्रकाश परिणामों के साथ उत्साहित पर आधारित है। प्रतिदीप्ति anisotropy 1 समीकरण द्वारा परिभाषित किया गया है:
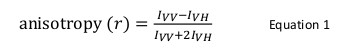
मैं वी.वी. और मैं VH रहे हैं, जहांखड़ी (वी.वी.) और क्षैतिज (VH) के प्रतिदीप्ति तीव्रता उत्सर्जन ध्रुवीकरण जब नमूना उत्साहित है खड़ी साथ प्रकाश 12 ध्रुवीकरण। प्रतिदीप्ति anisotropy कारक है कि fluorophore की बारी-बारी से प्रसार की दर को प्रभावित करने के लिए संवेदनशील है और इस तरह के तापमान, समाधान की चिपचिपाहट और fluorophore का स्पष्ट आणविक आकार पर निर्भर करता है। एक प्रोटीन एक fluorophore बढ़ जाती युक्त जब यह एक और प्रोटीन और इस तरह के परिवर्तन के साथ सूचना का आदान प्रदान के स्पष्ट आकार तो anisotropy में बदलाव के रूप में मूल्यांकन किया जा सकता है। अधिक विशेष रूप से, एक fluorophore है कि इसके फ्लोरोसेंट जीवन भर के लिए समाधान के सापेक्ष में धीरे-धीरे घूमता है एक अपेक्षाकृत बड़े anisotropy प्रदर्शन करेंगे एक बड़ा मैं वी.वी. मूल्य और छोटे मैं VH मूल्य नहीं है और इसलिए होगा। Fluorophores कि तेजी से अपने फ्लोरोसेंट जीवन भर के सापेक्ष हुई बात के लिए, मैं वी.वी. और मैं VH समान हो जाएगा और उनके anisotropy मूल्य छोटा हो जाएगा 12 </sup> (चित्रा 1)। इसके अलावा, शोर माप के लिए एक अच्छा संकेत anisotropy के लिए, यह आवश्यक है ब्याज की अणु की बारी-बारी से सह-संबंध समय के लिए इसी तरह एक प्रतिदीप्ति जीवन भर के साथ एक fluorophore के लिए है। अन्यथा, यह सही मुक्त प्रोटीन के बीच और कहा कि परिसर में anisotropy में अंतर रिकॉर्ड करने के लिए संभव नहीं है। उदाहरण के लिए, एक जीवन भर के 100 दा का एक कम आणविक भार परिसर से जुड़ी ऐसी fluorescein या rhodamine के रूप में 4 NSEC के पास से एक फ्लोरोसेंट जांच की anisotropy 0.05 है। 160 केडीए 0.29 करने के लिए अपने anisotropy मूल्य में वृद्धि होगी एक अणु के लिए बाइंडिंग; एक अंतर यह है कि सही ढंग से मापा जा सकता है। इसके विपरीत, एक ही फ्लोरोसेंट एक बाध्यकारी प्रतिक्रिया आणविक आकार में वृद्धि जिसका 1,000 केडीए के लिए 65 से भिन्न होता है में शामिल जांच केवल 0.3 करने के लिए 0.28 की एक anisotropy परिवर्तन है, जो बहुत छोटा सही मापा जा रहा है में परिणाम होगा। इस परिदृश्य में, 400 nsec का एक जीवन भर के साथ एक अधिक उपयुक्त जांच 12 होगा।
<pवर्ग = "jove_content">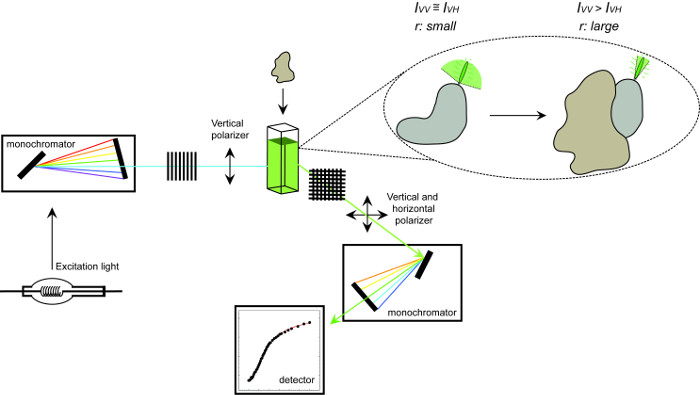
चित्रा 1. प्रतिदीप्ति anisotropy और प्रक्रिया को मापने के लिए इस्तेमाल किया उपकरणों की योजनाबद्ध प्रतिनिधित्व। एक प्रोटीन, प्रोटीन बातचीत प्रयोग मापने प्रतिदीप्ति anisotropy प्रदर्शन करने के लिए इस्तेमाल किया उपकरणों की योजनाबद्ध प्रतिनिधित्व। Fluorophores है कि तेजी से प्रदर्शन छोटे anisotropy कि एक बातचीत साथी के लिए बाध्य पर बढ़ जाती हुई। यह आंकड़ा का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।
प्रतिदीप्ति अनुप्रयोगों के अणुओं का अध्ययन से किसी में एक fluorophore की उपस्थिति की आवश्यकता होती है। प्रोटीन, प्रोटीन बातचीत का अध्ययन करने के लिए वहाँ fluorophores के तीन प्रकार हैं: 1) नियासिन प्रोटीन में मौजूद अवशेषों, 2) रासायनिक जुड़ी fluorophores और 3) फ्लोरोसेंट संलयन भागीदारों के ऐसे हरी फ्लोरोसेंट प्रोटीन (GFP) और उसके Deriva के रूप मेंtives। सबसे अधिक प्रोटीन, एक बातचीत को मापने के लिए सबसे आसान तरीका है इस प्रकार इसी प्रतिदीप्ति स्पेक्ट्रा में परिवर्तन की निगरानी के द्वारा या tryptophan अवशेषों की प्रतिदीप्ति तीव्रता में परिवर्तन की निगरानी कर रहा है इसकी संरचना पर tryptophan अवशेषों की है। हालांकि, नियासिन अवशेषों विश्लेषण उलझी दोनों प्रोटीन में मौजूद हो सकता है। दूसरी ओर, एक fluorophore एक बातचीत के कारण फ्लोरोसेंट गुणों को बदलने के लिए उस पर या बाध्यकारी साइट के पास स्थित होने की जरूरत है और यह बातचीत के साथ ही हस्तक्षेप कर सकता है। यह विशेष ध्यान जब इस तरह के GFP के रूप में भारी fluorophores का उपयोग कर की जरूरत है। इन fluorophores से कोई बाध्यकारी अध्ययन के लिए इस्तेमाल किया जा सकता है यदि ऐसा है तो, शामिल प्रोटीन से एक के लिए बाह्य fluorophores लागू करने के लिए आवश्यक है। कई रासायनिक संश्लेषित fluorophores मौजूद हैं और covalently ऐसे एमाइन समूह (lysines या एन टर्मिनस के पक्ष श्रृंखला) और सिस्टीन में thiol समूहों के रूप में उनके प्रतिक्रियाशील समूहों के माध्यम से प्रोटीन के लिए संलग्न किया जा सकता है। एफआइसोथियोसाइनेट और succinimidyl एस्टर के साथ luorophore डेरिवेटिव एमाइड समूहों के साथ प्रतिक्रिया करते हुए iodoacetamide और maleimide thiol प्रतिक्रियाशील समूहों 13 हैं। सबसे आम प्रतिदीप्ति अनुप्रयोगों में इस्तेमाल रंगों fluorescein के डेरिवेटिव और rhodamine हरे रंग, coumarins, BODIPY fluorophores और एलेक्सा स्त्राव रंगों कर रहे हैं। व्यावसायिक रूप से उपलब्ध fluorophores और उनके उपयोग की एक विस्तृत सूची संदर्भों 14,15 में पाया जा सकता है। सफल लेबलिंग के लिए, प्रतिक्रियाशील समूह प्रोटीन की सतह पर उजागर किया जाना चाहिए, लेकिन प्रतिक्रियाशील कार्य समूहों को आम तौर पर polypeptides में मौजूद बड़ी संख्या के कारण यह साइट विशेष संशोधन पाने के लिए बहुत मुश्किल है। इस अध्ययन में ब्याज की प्रोटीन, SBDS, 5 मुक्त cysteines और 33 lysines कि कई साइट लेबलिंग में परिणाम हो सकता है शामिल हैं। गैर वर्दी लेबलिंग बाध्यकारी प्रभावित कर सकता है और डेटा विश्लेषण को मुश्किल के रूप में विभिन्न fluorophore अणुओं बाध्यकारी पर विभिन्न फ्लोरोसेंट तीव्रता संकेतों बटोर सकता है। ove करने के लिएइस समस्या rcome, हम फ्लैश fluorophore, 4 ', 5'-बीआईएस (1,3,2 dithioarsolan-2-YL) साइट प्रत्यक्ष लेबल करने के लिए fluorescein SBDS प्रोटीन का इस्तेमाल किया। यह एक मूल भाव अनुक्रम CCXXCC जहां एक्स किसी भी अमीनो सिस्टीन 16,17 के अलावा अन्य एसिड होता है से मिलकर फ्लैश-टैग के रूप में जानते में चार स्थान दिया गया है cysteines के लिए एक उच्च आत्मीयता के साथ एक arsenoxide डाई है। इस tetracysteine मूल भाव एक साथ प्रोटीन की समग्र गुना के विघटन को रोकने के लिए एक उपयुक्त लिंकर साथ एन या प्रोटीन की सी टर्मिनस जेनेटिक इंजीनियरिंग से जोड़ा है। फ्लैश डाई और फ्लैश-टैग से मिलकर जोड़ी मूल कोशिकाओं 17 में रहने वाले साइट विशेष लेबल प्रोटीन के लिए डिजाइन किया गया था, लेकिन यह भी इन विट्रो में शुद्ध प्रोटीन लेबल करने के लिए के रूप में यह यहाँ उदाहरण है इस्तेमाल किया जा सकता है। इसके अतिरिक्त, एंजाइमी रणनीतियों भी प्रोटीन 18 की साइट विशेष functionalization सक्षम करने के लिए विकसित किया गया है।
इस पांडुलिपि में हम प्रतिदीप्ति anisotropy एक की उपयोगिता का वर्णनSA उपकरण प्रोटीन, प्रोटीन बातचीत का अध्ययन करने के लिए। बंधन बाध्यकारी वक्र आकार के सरल निरीक्षण द्वारा मूल्यांकन किया जा सकता है, जबकि मात्रात्मक जानकारी प्रयोगात्मक डेटा के फिट से प्राप्त किया जा सकता है।
