세포는 끊임없이 서로 상호 작용 생체 거대 분자의 다수 포함되어 있습니다. 이 연관은 신호 전달, 유전자 발현 및 다른 이들 세포 이동의 조절에서 이들의 기능을 담당 세포 경로에 참여 착물을 일으킨다. 세포 내에서 발생하는 모든 단백질 – 단백질 상호 작용은 상호 작용 체라고하는 네트워크를 포함한다. 사카로 마이 세스 세레 비지에서의 단백질의 70 % 이상이 상호 작용 파트너 일을하는 것으로 나타났다. 셀의 상호 작용 체를 이해하고 그 기능은 복잡성과 생물의 다양성에 관련 정보를 제공합니다. 여러 방법은 단백질 – 단백질 상호 작용을 확인하고 특성화하기 위해 설명되었다. 효모 두 하이브리드 2, 단백질 단편 보완 분석 (3), 질량 분석 및 단백질 microarra에 결합 친 화성 정제 4로 넣어 방법을 통해 높은 다른YS는 5,6- 작용을 식별하기 위해 사용된다. 식별되면,이를 검증하기 위해 필요하며, 이는 경우에 따라 달라질 수있다. 일반적으로, 이들 실험은 유전자 결실 또는 단백질 중 하나의 과다 발현에 의해 상호 작용 쌍 등의 각 부재의 수준에서 작용 자체를 방해 포함하고 상기 다른 부재의 특성의 변화 나 함수를 찾고 세포 수준. 이어서, 생물 리 학적 기법 7은 분자 수준에서 단백질 – 단백질 상호 작용을 특성화하기 위해 사용된다. 열량 형광 분광법 정량적 및 기계적으로이를 설명하기 위해 사용되는 동안이를 위해, 단백질 복합체의 구조는 X 선 결정학, 핵 자기 공명 및 극저온 전자 현미경에 의해 결정된다.
본 연구에서는 형광의 이방성이는 GTPase EFL1 및 SBD 간의 상호 작용을 특성화하는 방법을 사용 하였다S 단백질. 이 단백질은 60S 리보솜 서브 유닛 (8)의 표면에서 진핵 개시 인자 (6)의 방출을 촉진하여 리보솜의 합성에 참여하고 있습니다. SBDS 단백질 EFL1는 구아노 신 다이 포스페이트 (10, 11)에 대한 그것의 친화도를 감소시키는 구아닌 뉴클레오타이드 교환 인자로서 Shwachman 증후군 다이아몬드-9 및 행위로 알려진 질환 돌연변이된다. SBDS의 질병 돌연변이는 EFL1과의 상호 작용을 폐지하고, 따라서 그것의 활성화를 방지 할 수 있습니다.
형광의 이방성은 일반적으로 단백질 – 펩타이드 또는 단백질 – 핵산 상호 작용을 연구하는 생물학 분야에 사용된다. 이것은 형광 물질이 부분적으로 편광 된 방사의 편광 결과 여기 있다는 원리에 기초한다. 형광 이방성은 수학 식 1에 의해 정의된다 :
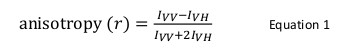
나는 절 내가 VH는 어디에수직 편광 된 광 (12)에 시료가 여기 될 때 수직 (VV) 및 가로 (VH)의 형광 발광 강도를 편광. 형광 이방성 형광의 회전 확산 속도에 영향을주는 요인에 민감하므로, 온도, 용액의 점도 및 형광 물질의 겉보기 분자 크기에 따라 달라진다. 또 다른 단백질과 같은 변화와 상호 작용할 때 형광 증가를 함유하는 단백질의 겉보기 크기이어서 이방성의 변화로 평가 될 수있다. 구체적으로는, 그것의 형광 수명을 용액 상대적으로 천천히 회전 형광 비교적 큰 이방성을 나타내는 것 때문에 큰 I VV의 값 I VH 작은 값을 가질 것이다. 자신의 형광 수명에 빠르게 상대 공중제비 형광, 나는 절 내가 VH는 유사합니다 및 이방성 값이 작은 것 (12) </sup> (그림 1). 또한, 노이즈 측정에 좋은 이방성 신호를 들면, 대상 분자의 회전 상관 시간 유사한 형광 수명을 가진 형광 물질이 필요하다. 그렇지 않으면 정확하게 복잡 프리 단백질과 그 이방성의 차이를 기록 할 수 없다. 예를 들어, 다 100의 저 분자량 화합물에 부착 같은 형광 또는 로다 민과 같은 4 NSEC 가까운 수명과 형광 프로브의 이방성은 0.05이다. kDa의 0.29로 그 이방성 값을 증가 (160)의 분자에 결합; 정확하게 측정 할 수있는 차이. 반면에, 그 증가 분자 크기가 변화 65 내지 1,000 kDa의 결합 반응에 관여 동일한 형광 프로브는 정확하게 측정하기에는 너무 작 0.3-0.28의 이방성이 변화 될 것이다. 이 시나리오에서 400 나노초의 수명이 프로브 (12)에 더 적합 할 것이다.
<p클래스 = "jove_content">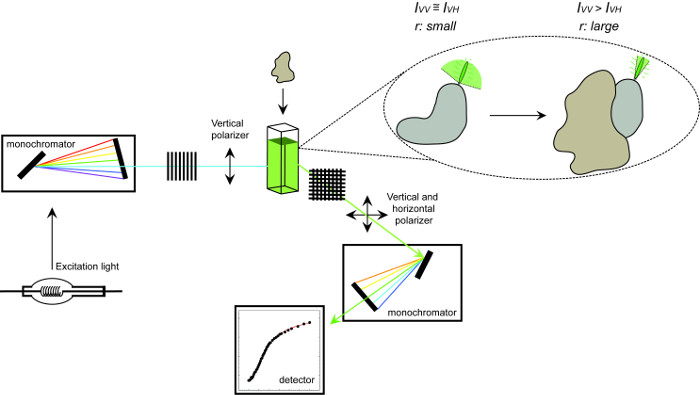
형광 이방성 절차를 측정하는 데 사용되는 장치의 개략도 1.도. 단백질 – 단백질 상호 작용의 실험 측정 형광 이방성을 수행하는 데 사용되는 장치의 개략도. 상호 작용 파트너의 결합에 증가 빠른 디스플레이 작은 이방성을 공중제비 형광. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
형광 응용 연구 분자 중 어느 하나의 형광의 존재를 필요로한다. 형광의 세 가지 유형이 단백질 – 단백질 상호 작용을 연구하는 방법 : 1) 단백질에 존재하는 트립토판 잔류 물, 2) 녹색 형광 단백질 (GFP)와 deriva 같은 화학적으로 부착 된 형광 3) 형광 융합 파트너tives. 대부분의 단백질은 그 구조의 상호 작용을 측정하는 것이 가장 간단한 방법 대응 형광 스펙트럼의 변화를 모니터링함으로써 또는 트립토판 잔기의 형광 강도의 변화를 감시함으로써이다 트립토판 잔기를 갖는다. 그러나, 트립토판 잔기의 분석을 복잡 단백질 모두에 존재할 수있다. 형광 인한 상호 작용을 형광의 속성을 변경하는 한편, 또는 결합 부위 근처에 위치해야하며 상호 자체를 방해 할 수있다. 같은 GFP와 같은 부피가 큰 형광체를 사용할 때 특별한주의가 필요합니다. 이 형광체 중에 결합 연구에 사용 할 수없는 경우는, 관여하는 단백질의 하나 외인성 형광체를 도입 할 필요가있다. 많은 화학적으로 합성 된 형광체는 존재 공유 같은 아민 기 (라이신 또는 N 말단의 측쇄) 및 시스테인의 티올기를 그들의 반응성기를 통해 단백질에 부착 될 수있다. 에프요오도 아세트 아미드 및 말레이 미드가 티올 – 반응성 기 (13) 동안 이소 티오 시아 네이트 및 숙신 이미 딜 에스테르로 luorophore 유도체 아미드 기와 반응. 형광 애플리케이션에서 가장 일반적으로 사용되는 염료는 플루오 레세 인 유도체 및 로다 민 그린 염료, 쿠마린, BODIPY 형광 단 알렉사 형석 염료이다. 상업적으로 입수 가능한 형광체와 그 사용의 자세한 목록은 참고 문헌 14, 15에서 볼 수있다. 성공적인 라벨링, 반응성 그룹은 단백질 표면에 노출되어야하지만 의한 폴리펩티드 전형적으로 존재하는 반응성 관능기의 다수는 부위 특이 적 변형을 얻는 것은 매우 어렵다. 이 연구에 대한 관심의 단백질, SBDS, 여러 사이트 표시 될 수 있습니다 5 무료 시스테인, 33 라이신이 포함되어 있습니다. 불균일 한 표시는 결합에 영향을 미칠 수있는 다른 형광 분자가 결합에 따라 상이한 형광 강도 신호를 유도 할 수있다으로 데이터 분석을 복잡하게한다. 비켜하려면이 문제를 rcome 우리는 플래시 형광, 4 '부위 – 직접 라벨, 5'- 비스 (1,3,2 dithioarsolan -2- 일) – 플루오 레세 SBDS 단백질을 사용 하였다. 이 모티프는 X 시스테인 (16, 17) 이외의 아미노산 서열로 이루어진 플래시 CCXXCC 태그로 알려진 네 개의 이격 시스테인에 대한 높은 친 화성을 갖는 arsenoxide 염료이다. 이 tetracysteine 모티브 함께 단백질의 전체 스크롤의 중단을 방지하기 위해 적당한 링커로 유전 공학에 의해 N- 또는 단백질의 C 말단에 부가된다. 플래시 염료 및 플래시 태그 이루어지는 쌍은 원래 셀 17 거주 지역 사이트 별 라벨 단백질 설계되었지만, 또한 여기에 열거 된 바와 같이 시험 관내에서 정제 된 단백질의 라벨을 사용할 수있다. 또한, 효소 적 전략은 단백질 (18)의 부위 특이 적 작용을 가능하게하기 위해 개발되어왔다.
이 논문에서는 형광 이방성 (A)의 유용성을 설명SA 도구는 단백질 – 단백질 상호 작용을 연구합니다. 정량적 인 정보는 실험 데이터의 피트로부터 얻어 질 수있는 반면 결합은 결합 곡선 형상의 간단한 검사에 의해 평가 될 수있다.
