Клетки содержат множество биомакромолекулах, которые постоянно взаимодействуют друг с другом. Эта ассоциация рождает комплексы, которые участвуют в клеточных путей, ответственных за их функционирование в сигнальной трансдукции, регуляции экспрессии генов и миграции клеток среди других. Все белок-белковых взаимодействий, которые происходят в клетке составляют сеть, известную как интерактома. В Saccharomyces CEREVISIAE было показано более чем 70% от его белков , чтобы иметь взаимодействующих партнеров 1. Понимание интерактома клетки и их функции предоставляют соответствующую информацию о сложности и разнообразии живых организмов. Несколько методик были описаны для идентификации и характеристики белок-белковых взаимодействий. Различные высоко через положить методы , такие как дрожжи дигибридная 2, белок-фрагмент комплементационных анализов 3, 4 аффинной очистки в сочетании с масс – спектрометрии и белка microarraYS используются для идентификации взаимодействия 5,6. После идентификации, необходимо проверить его, и это может варьироваться в зависимости от случая к случаю. Как правило, эти эксперименты связаны с срыве самого взаимодействия на уровне отдельных членов пары взаимодействия, например, путем делеции гена или избыточная экспрессия одного из белков, а затем ищет изменения в свойствах или функции другого члена в то на клеточном уровне. Впоследствии, биофизические методы 7 используются для характеристики белок-белковых взаимодействий на молекулярном уровне. С этой целью, структура белковых комплексов определяются с помощью рентгеновской кристаллографии, ядерного магнитного резонанса и крио-электронной микроскопии в то время как калориметрии и флуоресцентной спектроскопии используются для количественной и механистически описания.
В этой работе, анизотропии флуоресценции использовали в качестве методики для характеристики взаимодействия между ГТФ EFL1 и SBDS белок. Эти белки участвуют в синтезе рибосом путем содействия высвобождение эукариотических фактора инициации 6 от поверхности 60S рибосомальной субъединицей 8. Белок SBDS мутирует в заболевание , известное как Shwachman-Даймонд синдром 9 и действует как фактор обмена гуанина нуклеотиддля EFL1 уменьшая его сродство к гуанозиндифосфат 10,11. мутации болезни в SBDS отменить взаимодействие с EFL1 и тем самым предотвратить его активацию.
Флуоресцентная анизотропия обычно используется в биологических приложениях для изучения белок-пептид или взаимодействий белок-нуклеиновых кислот. Он основан на принципе, что флуорофор возбуждаемых с поляризованными светом приводит частично поляризованного излучения. Анизотропии флуоресценции определяется уравнением 1:
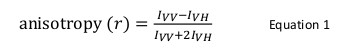
где я VV , и я В.Х. являютсяинтенсивности флуоресценции вертикально (VV) и по горизонтали (VH) поляризован излучение , когда образец возбуждается с вертикально поляризованный свет 12. Флуоресцентная анизотропия чувствительна к факторам, которые влияют на скорость вращательной диффузии флуорофора и, таким образом, зависит от температуры, вязкости раствора и кажущегося размера молекул флуорофора. Видимый размер белка, содержащего флуорофор увеличивается, когда он взаимодействует с другим белком, и такое изменение может быть оценена как изменение анизотропии. Более конкретно, флуорофор , который вращается медленно в растворе по отношению к его жизни флуоресцентного будет иметь большое значение , я VV и малое значение I V H , и , следовательно , будет демонстрировать относительно большую анизотропию. Для флуорофоров , что ОБРУШЬТЕСЬ быстро по сравнению с их флуоресцентным жизни, я VV , и я В.Х. будут подобны и их анизотропии значение будет небольшим 12 </sup> (Рисунок 1). Кроме того, для хорошей анизотропии сигнал измерения уровня шума, необходимо иметь флуорофор с времени жизни флуоресценции, подобной времени вращательной корреляции интерес молекулы. В противном случае, не представляется возможным точно регистрировать разницу в анизотропии между свободным белком и что в комплексе. Например, анизотропия флуоресцентного зонда с временем жизни около 4 наносекунд, такие как флуоресцеин или родамин, прикрепленных к низкомолекулярного соединения молекулярного 100 дальтон составляет 0,05. Связывание с молекулой 160 кДа увеличит его анизотропии значение 0,29; различие, которое может быть точно измерена. В противоположность этому, тот же флуоресцентного зонда участвуют в реакции связывания, чей рост размера молекул варьируется от 65 до 1000 кДа, приведет лишь к изменению анизотропии 0,28 до 0,3, который слишком мал, чтобы быть точно измерена. В этом случае зонд со временем жизни 400 нсек был бы более подходящим 12.
<pкласс = "jove_content">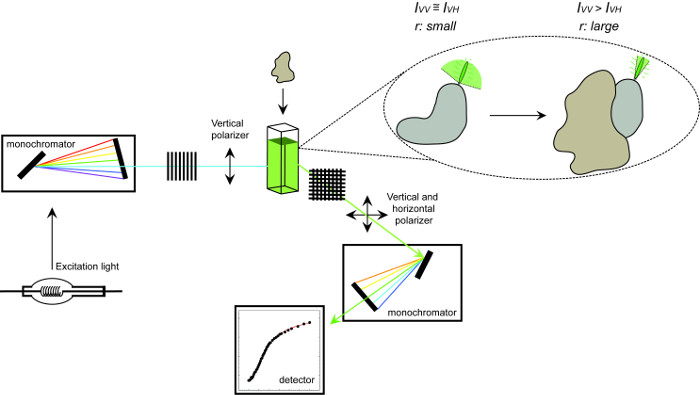
Рисунок 1. Схематическое изображение оборудования , используемого для измерения анизотропии флуоресценции и процедуры. Схематическое изображение оборудования , используемого для выполнения измерительный белок-белок эксперимент взаимодействия анизотропии флуоресценции. Флуорофоры , что барабанные быстрый дисплей небольшой анизотропии , которая увеличивает на связывание с партнером взаимодействия. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Флуоресцентные системы требуют присутствия флуорофор в любой из молекул изученных. Для изучения белок-белковых взаимодействий, существуют три типа флуорофоров: 1) Триптофан остатки, присутствующие в белках, 2) химически присоединенные флуорофоры и 3) флюоресцентные партнеры слитые, такие как зеленый флуоресцентный белок (GFP) и его Derivaставители. Большинство белков имеют остатки триптофана на его структуру, таким образом, самый простой способ измерить взаимодействие является путем мониторинга изменений в соответствующих спектров флуоресценции или путем наблюдения за изменениями в интенсивности флуоресценции остатков триптофана. Тем не менее, триптофан остатки могут присутствовать в обоих белках, усложняющих анализ. С другой стороны, для флуорофора, чтобы изменить его флуоресцентные свойства из-за взаимодействия он должен быть расположен на или вблизи сайта связывания, и это может помешать само взаимодействие. Это требует особого внимания при использовании громоздких флуорофоры, такие как GFP. Если ни один из этих флуорофоров не может быть использован для исследований связывания необходимо, а затем, чтобы ввести примесные флуорофоры к одному из белков, участвующих. Многие химически синтезированные флуорофоры существовать и могут быть ковалентно связаны с белками через их реакционноспособные группы, такие как аминные группы (боковой цепи лизина или N-конца) и тиоловых групп в цистеин. Fпроизводные luorophore с изотиоцианатных и сукцинимидиловых эфиры реагируют с амидной группы в то время как йодацетамида и малеимидная являются тиол-реактивные группы 13. Наиболее распространенные красители, используемые в приложениях флуоресцентных являются производными флуоресцеина и родамина зеленые красители, кумарины, BODIPY флуорофоры и Alexa Fluor красители. Подробный перечень коммерчески доступных флуорофоров и их использование можно найти в ссылках 14,15. Для успешной маркировки, реактивная группа должна быть открыта на поверхности белка, но из-за большого количества химически активных функциональных групп, обычно присутствующих в полипептидах очень трудно получить модификации конкретного участка. Белок интерес в данном исследовании, SBDS, содержит 5 свободных цистеина и 33 лизинов, которые могут привести к многократной маркировки сайта. Неравномерный маркировка может повлиять на связывании и усложнит анализ данных, как разные молекулы флуорофором могут вызывать различные сигналы интенсивности флуоресценции при связывании. Для Овеrcome эту проблему, мы использовали флуорофора âñïûøêè, 4 ', 5'-бис (1,3,2 dithioarsolan-2-ил) флуоресцеина сайт-прямой этикетке белок SBDS. Это arsenoxide краситель с высоким сродством к четырем разнесенных цистеина в мотиве знают , как ФЛЭШ-тег , состоящий из CCXXCC последовательности , где X обозначает любую аминокислоту, кроме цистеина 16,17. Этот tetracysteine мотив добавлен к N- или С-конце белка методом генной инженерии вместе с соответствующим линкером для предотвращения нарушения общей складки белка. Пара , состоящая из красителя и ФЛЭШ ФЛЭШ-таг был первоначально разработан для сайт-специфических белков меток в живых клетках 17 , но он также может быть использован для обозначения очищенных белков в пробирке , как это иллюстрируется здесь. Кроме того, ферментативные стратегии были также разработаны для того, чтобы сайт-специфической функционализации белков 18.
В этой рукописи мы опишем полезность флуоресцентной анизотропии аса инструмент для изучения белок-белковых взаимодействий. Связывание может быть оценена с помощью простого осмотра связывающей формы кривой в то время как количественная информация может быть получена из подгонки экспериментальных данных.
