Celler innehåller en mångfald av biomakromolekyler som ständigt interagerar med varandra. Denna förening ger upphov till komplex som deltar i de cellulära vägar som ansvarar för deras funktion i signalöverföring, reglering av genuttryck och cell migration bland andra. Alla protein-proteininteraktioner som sker i en cell innefattar ett nätverk som kallas interactome. I Saccharomyces cerevisiae har visats mer än 70% av dess proteiner för att ha interagerande partners 1. Förstå interactome av en cell och deras funktioner ger relevant information på komplexiteten och mångfalden av levande organismer. Flera metoder har beskrivits för att identifiera och karakterisera protein-proteininteraktioner. Olika hög genomströmningshastighet metoder såsom jäst två-hybrid-2, proteinfragmentkompletteringsanalyser 3, affinitetsrening 4 kopplad till masspektrometri och protein microarrays används för att identifiera en interaktion 5,6. När detta har skett, är det nödvändigt att bekräfta det och kan variera från fall till fall. Normalt är dessa experiment involverar att störa interaktionen sig på samma nivå som de enskilda medlemmarna i samspelet paret, t ex genom gendeletion eller överuttryck av ett av proteinerna, och sedan letar efter förändringar i egenskaper eller funktion hos andra medlem på cellnivå. Därefter biofysiska tekniker 7 används för att karakterisera interaktion protein-protein på molekylnivå. För detta ändamål, är strukturen av proteinkomplex bestämdes genom röntgenkristallografi, kärnmagnetisk resonans och Cryo-elektronmikroskopi medan kalorimetri och fluorescensspektroskopi används för att kvantitativt och mekanistiskt beskriva dem.
I detta arbete, var fluorescensanisotropi används som en teknik för att karakterisera interaktionen mellan GTPas EFL1 och SBDS-proteinet. Dessa proteiner deltar i syntesen av ribosomer genom att främja frisättningen av eukaryota inledande faktor 6 från ytan av 60S ribosomala subenheten 8. Den SBD-protein är muterat i en sjukdom som är känd som den Shwachman-Diamond syndrom 9 och fungerar som en guaninnukleotidutbytesfaktor för EFL1 minska dess affinitet för Guanosindifosfat 10,11. Sjukdoms mutationer i SBD avskaffa interaktionen med EFL1 och därmed förhindra dess aktivering.
Fluorescensanisotropi används allmänt i biologiska tillämpningar för att studera protein-peptid eller protein-nukleinsyra-interaktioner. Den bygger på principen att en fluorofor exciteras med polariserat ljus resulterar i en delvis polariserad emission. Fluorescensanisotropi definieras av ekvation 1:
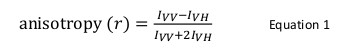
där jag Vv och jag VH ärfluorescensintensiteter av vertikalt (VV) och horisontellt (VH) polariserad emission när provet exciteras med vertikalt polariserat ljus 12. Fluorescensanisotropi är känslig för faktorer som påverkar graden av rotations diffusion av fluoroforen och sålunda beror på temperaturen, viskositeten hos lösningen och den synbara molekylstorleken av fluoroforen. Den skenbara storleken av ett protein innehållande en fluorofor ökar när det interagerar med ett annat protein och en sådan förändring kan därefter utvärderas som en ändring i anisotropi. Mer specifikt kommer en fluorofor som roterar långsamt i lösning i förhållande till dess fluorescerande livslängd har en stor jag VV värde och liten jag VH värde och därför kommer att uppvisa en relativt stor anisotropi. För fluoroforer som tumlar snabbt i förhållande till deras fluorescerande livstid, jag vv och jag VH kommer att likna och deras anisotropi värdet kommer att vara liten 12 </sup> (Figur 1). Dessutom, för en god anisotropi signalbrusmätning, är det nödvändigt att ha en fluorofor med en fluorescenslivstid liknar rotationskorrelationstiden av molekylen av intresse. Annars är det inte möjligt att på ett korrekt skillnaden i anisotropi mellan fritt protein och att i komplexet. Till exempel, är anisotropin hos en fluorescerande sond med en livstid nära 4 nsek såsom fluorescein eller rodamin bundna till en lågmolekylär förening med 100 Da 0,05. Bindning till en molekyl av 160 kDa kommer att öka sin anisotropi värde till 0,29; en skillnad som kan mätas noggrant. Däremot kommer samma fluorescerande prob involverad i en bindningsreaktion, vars ökning i molekylstorleken varierar från 65 till 1.000 kDa endast resultera i en anisotropi förändring av 0,28 till 0,3, vilket är för liten för att mätas noggrant. I detta scenario skulle en sond med en livslängd på 400 ns vara lämpligare 12.
<pclass = "jove_content">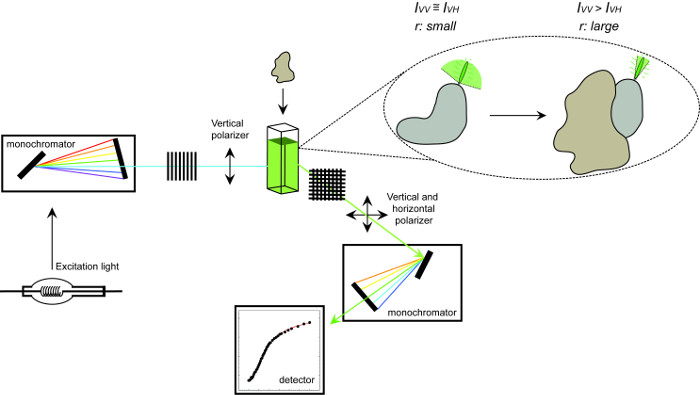
Figur 1. Schematisk representation av den utrustning som används för att mäta fluorescens anisotropi och proceduren. Schematisk representation av den utrustning som används för att utföra en protein-proteininteraktion experiment mätning av fluorescens anisotropi. Fluoroforer som tumlar snabbt display liten anisotropi som ökar vid bindning till en interaktion partner. Klicka här för att se en större version av denna siffra.
Fluorescensapplikationer kräver närvaro av en fluorofor i någon av molekylerna som studerats. För att studera protein-proteininteraktioner finns tre typer av fluoroforer: 1) de tryptofanrester närvarande i proteinerna, 2) kemiskt bundna fluoroforer och 3) fluorescerande fusionspartners såsom grönt fluorescerande protein (GFP) och dess Derivahavare. De flesta proteiner har tryptofanrester på dess struktur, således det enklaste sättet att mäta en interaktion är genom att övervaka förändringarna i den motsvarande fluorescensspektra eller genom övervakning av förändringar i fluorescensintensiteten hos de tryptofanrester. Emellertid kan tryptofangrupper vara närvarande i båda proteinerna komplicerar analysen. Å andra sidan, för en fluorofor att ändra sina fluorescerande egenskaper beroende på en interaktion den behöver för att vara placerad på eller i närheten av bindningsstället och det kan störa interaktionen självt. Detta kräver särskild uppmärksamhet vid användning av skrymmande fluoroforer såsom GFP. Om ingen av dessa fluoroforer kan användas för bindningsstudier är det nödvändigt, då att införa yttre fluoroforer till ett av de proteiner som är inblandade. Många kemiskt syntetiserade fluoroforer existerar och kan fästas kovalent till proteiner via deras reaktiva grupper såsom amingrupper (sidokedjan i lysiner eller N-terminalen) och tiolgrupperna i cystein. Fluorophore derivat med isotiocyanat och succinimidylestrar reagerar med amidgrupper medan jodacetamid och maleimid är tiol-reaktiva grupper 13. De vanligaste färgämnen som används i fluorescensapplikationer är derivat av fluorescein och rodamin gröna färgämnen, kumariner, BODIPY fluoroforer och Alexa Fluor färgämnen. En detaljerad lista över kommersiellt tillgängliga fluoroforer och deras användning kan hittas i referenser 14,15. För framgångsrik märkning, måste den reaktiva gruppen att exponeras på ytan av proteinet, men på grund av det stora antalet reaktiva funktionella grupper som vanligen är närvarande i polypeptider är det mycket svårt att få platsspecifik modifiering. Proteinet av intresse i denna studie, SBD innehåller 5 fria cysteiner och 33 lysiner som kan resultera i flera plats märkning. Icke-enhetlig märkning kan påverka bindningen och kommer att försvåra dataanalys som olika fluoroforen molekyler kan framkalla olika fluorescerande intensitetssignaler vid bindning. att overcome detta problem använde vi Flash fluorofor, 4 ', 5'-bis (1,3,2 dithioarsolan-2-yl) fluorescein till plats direkt etikett SBD-protein. Detta är en arsenoxide färgämne med en hög affinitet för fyra åtskilda cysteiner i ett motiv vet som Flash-tag som består av sekvensen CCXXCC där X är vilken aminosyra som helst annan än cystein 16,17. Denna tetracysteine motiv sättes till N- eller C-terminalen av proteinet med genteknik tillsammans med en lämplig linker för att förhindra störningar av den totala vikningen av proteinet. Paret som består av flash-färgämne och FLASH-tagg var ursprungligen avsedd att platsspecifika etikett proteiner i levande celler 17 men det kan också användas för att märka renade proteiner in vitro såsom den exemplifieras här. Dessutom har enzymatiska strategier också utvecklats för att göra det möjligt för platsspecifik funktionalisering av proteiner 18.
I detta manuskript beskriver vi nyttan av fluorescens anisotropi ensa verktyg för att studera protein-proteininteraktioner. kan bedömas genom enkel inspektion av bindande kurvform bindande medan kvantitativ information kan erhållas från passningen av experimentella data.
