Interaktion mellem molekyler er grundlaget for naturen. Derfor forskere i mange områder af grundforskning og anvendt forskning forsøge at forstå de grundlæggende principper om molekylære interaktioner af forskellig art. Individuel Thermophoresis (MST) gør det muligt for forskere at udføre hurtig, præcis, omkostningseffektiv, og kvalitet-kontrollerede karakterisering af molekylære interaktioner i opløsning, med frit valg af buffere. Der er allerede mere end 1.000 publikationer MST, fra 2016 alene, der beskriver forskellige former for analyser, herunder bibliotekets screeninger, bindende event valideringer, konkurrence analyser og eksperimenter med flere bindende partnere 1-8. Generelt MST tillader studiet af de klassiske bindende parametre, såsom bindingsaffinitet (pM til mM), støkiometri og termodynamik, af enhver form for molekylær interaktion. En stor fordel ved MST er evnen til at studere bindingsbegivenheder uafhængigt af størrelsen af de interaktionspartnere. Selv udfordrende interaktioner mellem små nucleinsyre aptamerer (15-30 nt) og mål såsom små molekyler, lægemidler, antibiotika eller metabolitter kan kvantificeres.
Aktuelle state-of-the-art teknologier til at karakterisere aptamer målgruppen interaktioner er enten lab-intens og meget komplekse eller undlader at kvantificere aptamer-lille molekyle interaktioner 9,10. Surface (SPR) -baserede assays 11,12 og virkelig label-fri kalorimetriske tilgange, såsom Isotermisk Titrering kalorimetri (ITC) 13-15, isokratisk eluering 16, ligevægt fi ltrering 17,18, in-line sondering 19, gel- skift analyser, stopped- fl ow fl uorescence spektroskopi 20,21, fluorescens anisotropi (FA) 22,23, single-molekyle fl uorescence billedbehandling 24,25, og Bio-lags interferometri (BLI) 26 er også enten upræcise eller uforenelig med aptamer-lille molekyle interaktioner. Andre principal spørgsmål af disse metoder er lav følsomhed, højt forbrug prøve, immobilisering, masse begrænsninger transport på overflader, og / eller buffer restriktioner. Kun få af disse teknologier giver integrerede styringer til sammenlægning og adsorption effekter.
MST er et effektivt redskab for forskerne at overvinde denne begrænsning for at undersøge samspillet mellem aptamerer og små molekyler 27-29, samt andre mål, såsom proteiner 30-33. Teknologien bygger på flytning af molekyler gennem temperaturgradienter. Denne rettet bevægelse, kaldet "thermophoresis," afhænger af størrelse, ladning, og hydrering skallen af molekylet 34,35. Bindingen af en ligand til molekylet vil direkte ændre mindst en af disse parametre, hvilket resulterer i en ændret thermophoretic mobilitet. Ligander med små størrelser kan ikke have en betydelig indvirkning i form af størrelse ændring fra den ubundne til den bundne tilstand, men de kan have dr amatic virkninger på hydrering skal og / eller beregning. Ændringerne i thermophoretic bevægelse af molekyler efter interaktioner med bindingspartneren muliggør kvantificering af grundlæggende bindingsparametre 2,7,34,36,37.
Som afbildet i figur 1A, MST anordning består af en infrarød laser fokuseres på prøven inden for de glaskapillarer anvendelse af de samme optikker som for fluorescens detektion. Kan overvåges thermophoretic bevægelse af proteiner via den iboende fl uorescence af tryptophaner 6 eller af en fluorescens-mærket interaktion partner 3,8, mens laseren etablerer en temperaturgradient (AT på 2-6 ° C). Den resulterende temperaturforskel i rummet, AT, fører til udtømning eller akkumulering af molekyler i området for forhøjet temperatur, som kan kvantificeres ved Soret fi cient (S T):
g "/>
c varmt repræsenterer koncentrationen i den opvarmede region, og C Kold er koncentrationen i den indledende koldt region.
Som vist i figur 1B, en typisk MST eksperimentresultaterne i en MST bevægelse profil (tidssporet), der består af forskellige faser, som kan adskilles ved deres respektive tidsplaner. Den indledende fluorescens måles i de første 5 s i fravær af temperaturgradienten at definere den præcise start fluorescens og for at kontrollere for fotoblegning eller photoenhancement. Temperaturen Jump (T-jump) repræsenterer den fase, hvor fluorescens ændringer før thermophoretic bevægelse. Denne indledende fald i fluorescens afhænger varme-afhængige ændringer i fl uorophore kvantumudbytte. Den thermophoresis fase følger, hvor fluorescens falder (eller stiger) på grund af den thermophoretic bevægelse af molekylerne indtil steady-state fordelingen er nået.Kan observeres den omvendte TJump og samtidig tilbagediffusion af fl uorescent molekyler som vist i figur 1 B, efter at laseren er slukket. For at få adgang grundlæggende bindende parametre, er forskellige molære forhold af interaktionspartnere analyseret og sammenlignet. Typisk er 16 forskellige forhold undersøgt i en MST eksperiment, hvorimod den optiske synlige molekyle holdes konstant og leveres med en stigende mængde af umærket ligand. Samspillet mellem de to bindende partnere inducerer ændringer i thermophoresis, og dermed i det normaliserede fl uorescence, F norm, der beregnes som følger:
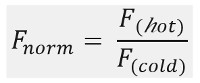
F varmt og F koldt repræsenterer gennemsnit fl uorescence intensiteter på defineret tidspunkter af MST spor. Bindingsaffiniteter (K d eller EC 50 værdier) kan beregnes ved Curve fitting (figur 1C).
Samlet set MST er et stærkt værktøj til at studere molekylære interaktioner af nogen art. Dette håndskrift tilbyder en protokol til at karakterisere udfordrende samspil mellem det lille molekyle adenosin trifosfat (ATP, 0,5 kDa) og 25-nt kort ssDNA aptamer DH25.42 (7,9 kDa). I løbet af manuskriptet, er bindingsstedet for aptameren på ATP-molekylet kortlagt ned til adenin gruppe af ATP.
