Взаимодействие между молекулами является основой природы. Таким образом, ученые во многих областях фундаментальных и прикладных исследований пытаются понять основные принципы молекулярных взаимодействий различных видов. Microscale Термофорез (MST) позволяет ученым осуществлять быстрый, точный, экономичный, и контроль качества характеристику молекулярных взаимодействий в растворе, со свободным выбором буферов. Есть уже более 1000 публикаций с использованием MST, с 2016 года в одиночку, описывая различные виды анализов, в том числе библиотеки скринингов, связывание валидаций событий, анализы конкуренции, а также эксперименты с несколькими партнерами связывания 1-8. В общем, МСТ позволяет исследовать классических параметров связывания, такие как аффинность связывания (стр.м.в мМ), стехиометрии и термодинамики, любого вида молекулярного взаимодействия. Большим преимуществом MST является возможность изучения связывания событий независимо от размера партнеров взаимодействия. Даже чалlenging взаимодействия между малыми аптамеров нуклеиновых кислот (15-30 нт) и задачи, такие как небольшие молекулы, лекарственные средства, антибиотики или метаболиты могут быть определены количественно.
Текущее состояние-оф-современные технологии для характеристики аптамеров-мишени взаимодействия либо лабораторного интенсивной и очень сложной или не в состоянии количественно аптамеров-малая молекула взаимодействиях 9,10. Поверхностного плазмонного резонанса (SPR) основе анализов 11,12 и действительно этикетки свободные калориметрических подходы, такие как изотермической калориметрии (титрования ITC), 13-15 изократическим элюирования 16, равновесному инфильтрации 17,18, рядный зондирования 19, гелеобразные анализах сдвига, stopped- потока флуоресценции спектроскопии 20,21, анизотропии флуоресценции (FA) 22,23, одной молекулы флуоресценции изображений 24,25 и Био-слой интерферометрии (BLI) 26 также являются либо неточными или несовместим с аптамеров-небольшой молекулы взаимодействия. Другое principaл вопросы этих методов низкая чувствительность, высокое потребление пробы, иммобилизация, ограничение массовых перевозок на поверхностях, и / или ограничения буфера. Лишь немногие из этих технологий обеспечивают интегрированные средства управления для агрегации и адсорбции эффектов.
MST представляет собой мощный инструмент для ученых , чтобы преодолеть это ограничение для изучения взаимодействия между аптамеров и малыми молекулами 27-29, а также другие цели , такие как белки 30-33. Технология опирается на движение молекул через температурных градиентов. Это направленное движение, называемое "термофорезу," зависит от размера, заряда и гидратной оболочки молекулы 34,35. Связывание лиганда с молекулой будет непосредственно изменять, по меньшей мере один из этих параметров, в результате чего в измененном термофоретического мобильности. Лиганды с малыми размерами не могут иметь значительное влияние с точки зрения изменения размера от несвязанного в связанном состоянии, но они могут иметь Др amatic воздействие на оболочку гидратации и / или заряда. Изменения в термофоретического движение молекул после взаимодействия с партнером по связыванию позволяет количественно оценить основные параметров связывания 2,7,34,36,37.
Как показано на рисунке 1А, устройство МСТ состоит из инфракрасного лазера фокусировалось на образец в стеклянных капиллярах с использованием тех же оптических систем для флуоресцентной детекции. Термофоретическая движение белков с помощью собственной флуоресценции триптофанов 6 или флуоресцентно меченого партнерского взаимодействия 3,8 можно отслеживать в то время как лазер создает температурный градиент (& Delta ; t 2-6 ° С). Полученная разность температур в пространстве, & Delta ; t, приводит к истощению или накопление молекул в области повышенной температуры, которая может быть определена количественно с помощью Соре коэффици- циент (С Т):
г "/>
С горячей представляет собой концентрацию в нагретой области, и с холодом концентрация в начальной холодной области.
Как показано на фигуре 1В, типичный результаты эксперимента MST в профиле движения MST (время трассировки), состоящий из различных этапов, которые могут быть разделены с помощью их соответствующих временных рамок. Первоначальная измеряют флуоресценцию в первые 5 секунд в отсутствие градиента температуры, чтобы определить точную начальную флуоресценцию и для проверки наличия фотообесцвечиванию или photoenhancement. Скачок температуры (T-Jump) представляет собой фазу, в которой изменения флуоресценции до термофоретического движения. Это начальное снижение флуоресценции зависит от тепловых зависящих от изменений фл uorophore квантового выхода. Фаза термофорезу следует, в которых флуоресценция уменьшается (или увеличивается) в связи с термофоретического движения молекул вплоть до стационарного распределения достигается.Обратное TJump и сопутствующее назад диффузия фл uorescent молекул можно наблюдать , как показано на рисунке 1В после того, как лазер выключен. Для того, чтобы получить доступ к основным параметрам связывания, различные молярные отношения партнеров взаимодействия анализируются и сравниваются. Как правило, 16 различных соотношений изучаются в одном эксперименте MST, в то время как оптический видимый молекулы поддерживается постоянным и поставляется с увеличением количества немеченого лиганда. Взаимодействие между этими двумя связывающими партнерами вызывает изменения в ТФ, и , таким образом , в нормированной флуоресценции, F норма, которая рассчитывается следующим образом :
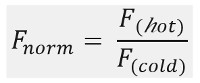
F горячей и холодной F представляют собой усредненные интенсивности uorescence Fl на де определены моменты времени следов MST. Аффинности связывания (K D или 50 значений EC) можно рассчитать по CURVе фитинга (рис 1C).
В целом, MST является мощным инструментом для изучения молекулярных взаимодействий любого рода. Эта рукопись предлагает протокол характеризовать сложные взаимодействия между молекулой небольшой аденозинтрифосфата (АТФ, 0,5 кДа) и 25-нт короткий оцДНК аптамеров DH25.42 (7,9 кДа). В течение рукописи, сайт связывания аптамеров на молекулы АТФ отображается вплоть до аденин группы АТФ.
