Estima-se que os genomas de mamíferos contêm cerca de 20.000 genes codificadores de proteínas. O splicing alternativo e modificações pós-tradução aumentam ainda mais o repertório de proteínas. As proteínas têm uma estrutura modular de um e muitas vezes conter vários domínios de interacção, que permitem o seu recrutamento para complexos proteicos diferentes e a sua participação em vários processos celulares 2. Um exemplo é a proteína multi-funcional chamado p120ctn. p120ctn é codificada pelo gene Ctnnd1 e consiste de um grande domínio central tatu repetição flanqueado por um N-terminal e uma região C-terminal. O domínio de tatu p120ctn se liga a um domínio justamembranar altamente conservado de caderinas clássicas, que estão envolvidos na adesão célula-célula, mas que também se liga ao repressor transcricional Kaiso. O domínio N-terminal de p120ctn interage com diferentes quinases, fosfatases, pequenas RhoGTPases, e associada a microtúbulos proteins 3. Curiosamente, como um resultado de splicing alternativo, isoformas p120ctn podem ser gerados a partir de quatro codões de iniciação alternativos 4. p120ctn isoforma 1A é a mais longa, uma vez que é traduzida a partir da maioria-5' do codão de iniciação e contém o segmento de comprimento completo N-terminal. Em p120ctn isoformas de 3 e 4, este segmento N-terminal é eliminado parcialmente e completamente, respectivamente. Compreendendo o papel preciso de proteínas (ou isoformas de proteas) e os seus domínios em diferentes funções celulares continua a ser um desafio.
Direccionamento de genes em mESCs permite o estudo da função de uma proteína através da deleco genica do gene correspondente e tem amplamente contribuiu para a identificação de genes e vias developmentally importantes e doenças relevantes. Esta descoberta em engenharia genica inversa foi o resultado de avanços em campos de isolamento MESC e direccionamento de genes devido à recombinação homóloga 5 </s-se>. A recombinação homóloga é um processo no qual fragmentos de ADN são trocadas entre duas porções nucleicos semelhantes ou idênticas depois de cadeia dupla (ds) quebras de ADN. Normalmente, RH é ineficiente porque quebras ADNcd são pouco frequentes. Recentemente, a eficiência do gene dirigida por homologia de segmentação pode ser aumentada usando nucleases específicas do local 6, 7, mas, infelizmente, estes são propensas a efeitos fora do alvo 8. Uma técnica mais fiável para permitir o direccionamento gico é RMCE, que é baseado em sistemas de recombinação específicos do local tais como Cre / loxP ou FLPe / Frt. LoxP e sequência DRF são encontrados no bacteriago P1 e Saccharomyces cerevisiae, respectivamente, e consistem de 34 pb, incluindo uma sequência de pb assimétrica 8 que determina a orientação do local. Por outro lado, a orientação de, por exemplo, dois sítios loxP no interior de um trecho de ADN irá determinar se o ADN floxed torna-se excisado ou inversed após recombinação mediada por Cre 9. Além disso, Cre também pode induzir uma translocação se dois locais estão localizados em diferentes cromossomas. RMCE leva vantagem de locais de recombinação heteroespecíficos que não reagem de forma cruzada e que são incorporados no locus genómico. Na presença de um plasmídeo dador que contém um fragmento de ADN flanqueada pelos mesmos locais heteroespecíficos, a recombinase irá inserir este fragmento de ADN no locus genómico RMCE-compatíveis, porque de translocação duplo simultâneo (Figura 1). Aqui, os clones única correctamente RMCE orientadas podem tornar de resistência a drogas, graças a um promotor no vector de entrada que restaura um "preso", gene de resistência à Neomicina promotor-menos (Neo R) presente no genoma R26 das células de encaixe (Figura 1) 10, 11. Isto resulta numa eficiência muito alta segmentação, muitas vezes perto de 100% 11, </ sup> 12. Em conclusão, a segmentação baseada RMCE é altamente eficiente e pode ser usada para a estrutura-funções estudos; no entanto, requer um local genómico pre-engenharia.
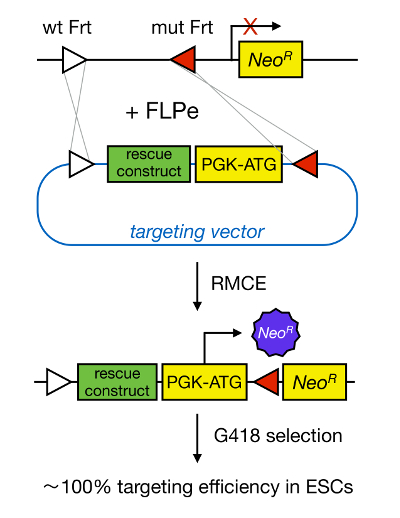
Figura 1. Representação esquemática da orientação para as mediada por RMCE. RMCE permite a troca de segmentos de ADN a partir de um vector de abordagem selectiva de entrada para um local genómico definido se tanto abrigar dois locais DRF heteroespecíficos (representados por triângulos brancos e vermelhos). Além disso, o locus genómico engenharia contém um promotor e um gene truncado de resistência à neomicina (NeoR). Ao fornecer um promotor e do codão de iniciação no fragmento de DNA de entrada, os eventos de recombinação única correcção restaurar a resistência a neomicina, resultando em altas eficiências de segmentação. Por favor clique aqui para ver uma versão maior do tsua figura.
engenharia genoma em mESCs permite a geração de ratinhos RMCE-compatível. Em 1981, dois grupos conseguiram capturar células pluripotentes a partir da massa celular interna (ICM) de blastocistos e em mantê-las em cultura 13, 14. mESCs são capazes de auto-renovação e diferenciação em todos os tipos de células embrionárias e adultos, incluindo a linhagem de células germinativas. Portanto, o direccionamento gico em mESCs permite estudos inverter-genéticas através do desenvolvimento de ratinhos KO constitutivos ou condicional (utilizando o sistema cre / loxP). No entanto, a forma clássica de isolar as células ES de ratinho é muito ineficiente. Várias melhorias importantes têm aumentado grandemente a taxa de sucesso para derivar linhas Mesc, incluindo o uso de um soro de substituição definidos (SR) de meio 15, alternando entre médio MESC contendo soro de bovino fetal e SR (FBS) a 16, e o uso de farmacocompostos lógicas tais como pluripotin ou 2i 17. Pluripotin, uma molécula pequena sintética, permite a propagação de mESCs num estado indiferenciado, na ausência do factor inibidor da leucemia (LIF) e fibroblastos de rato embrionários (MEFs) 18. Finalmente, foi demonstrado que mESCs pode ser isolado com um rendimento muito elevado (perto de 100%) quando um / FBS a meio protocolo alternância SR é combinado com LIF e pluripotin 19, 20. Estes protocolos permitem o isolamento eficiente de mESCs KO RMCE-compatível que pode subsequentemente ser utilizados para estudos estrutura-função.
Este documento descreve um método que permite que um para identificar os domínios de chave ou resíduos dentro de uma proteína que é responsável por processos celulares específicos. Para este fim, uma reserva de tecnologias avançadas que permitem o isolamento eficiente MESC, visando a montagem do vetor, e MESC segmentação era criard. Como tal, os grandes painéis com isoformas de proteas, mutantes de domínio, e efectores a jusante podem ser introduzidos em mESCs KO e podem ser avaliados pela sua capacidade para resgatar o fenótipo in vitro KO.
