It is estimated that mammalian genomes contain about 20,000 protein-coding genes. Alternative splicing and posttranslational modifications further increase the protein repertoire. Proteins have a modular structure1 and often contain multiple interaction domains, which allow their recruitment into different protein complexes and their participation in multiple cellular processes2. One example is the multi-functional protein called p120ctn. p120ctn is encoded by the Ctnnd1 gene and consists of a large central armadillo repeat domain flanked by an N-terminal and a C-terminal region. The armadillo domain of p120ctn binds to a highly conserved juxtamembrane domain of classical cadherins, which are involved in cell-cell adhesion, but it also binds to the transcriptional repressor Kaiso. The N-terminal domain of p120ctn interacts with different kinases, phosphatases, small RhoGTPases, and microtubule-associated proteins3. Interestingly, as a result of alternative splicing, p120ctn isoforms can be generated from four alternative start codons4. p120ctn isoform 1A is the longest, as it is translated from the most-5' start codon and contains the full-length N-terminal segment. In p120ctn isoforms 3 and 4, this N-terminal segment is deleted partially and completely, respectively. Understanding the precise role of proteins (or protein isoforms) and their domains in different cellular functions remains a challenge.
Gene targeting in mESCs enables the study of the function of a protein through the genetic deletion of the corresponding gene and has widely contributed to the identification of developmentally important and disease-relevant genes and pathways. This breakthrough in reverse genetics was the result of advances in the fields of mESC isolation and gene targeting due to homologous recombination5. Homologous recombination is a process in which DNA fragments are exchanged between two similar or identical nucleic moieties after double-stranded (ds) DNA breaks. Normally, HR is inefficient because dsDNA breaks are infrequent. Recently, the efficiency of homology-directed gene targeting could be increased using site-specific nucleases6,7, but unfortunately, these are prone to off-target effects8. A more reliable technique to enable gene targeting is RMCE, which is based on site-specific recombination systems such as Cre/loxP or FLPe/Frt. LoxP and Frt sequence are found in bacteriophage P1 and Saccharomyces cerevisiae, respectively, and consist of 34 bp, including an asymmetric 8 bp sequence that determines the orientation of the site. On the other hand, the orientation of, for instance, two loxP sites within a DNA stretch will determine whether the floxed DNA becomes excised or inversed upon Cre-mediated recombination9. Moreover, Cre can also induce a translocation if two sites are located on different chromosomes. RMCE takes advantage of heterospecific recombination sites that do not cross-react and that are embedded in a genomic locus. In the presence of a donor plasmid that contains a DNA fragment flanked by the same heterospecific sites, the recombinase will insert this DNA fragment into the RMCE-compatible genomic locus because of double-simultaneous translocation (Figure 1). Here, only correctly RMCE-targeted clones can render drug resistance thanks to a promoter on the incoming vector that restores a "trapped," promoter-less Neomycin resistance gene (NeoR) present in the R26 genome of the docking cells (Figure 1)10,11. This results in a very high targeting efficiency, often close to 100%11,12. In conclusion, RMCE-based targeting is highly efficient and can be used for structure-functions studies; however, it requires a pre-engineered genomic locus.
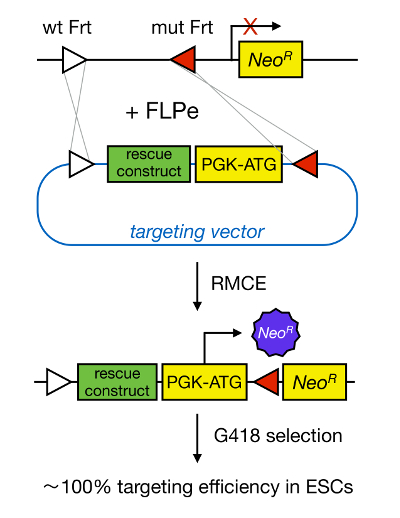
Figure 1. Schematic Representation of RMCE-mediated Targeting. RMCE allows for the exchange of DNA segments from an incoming targeting vector to a defined genomic locus if both harbor two heterospecific Frt sites (depicted by white and red triangles). In addition, the engineered genomic locus contains a promoterless and truncated neomycin-resistance (NeoR) gene. By providing a promoter and start codon in the incoming DNA fragment, only correct recombination events restore neomycin resistance, resulting in high targeting efficiencies. Please click here to view a larger version of this figure.
Genome engineering in mESCs allows for the generation of RMCE-compatible mice. In 1981, two groups succeeded in capturing pluripotent cells from the inner cell mass (ICM) of blastocysts and in maintaining them in culture13,14. mESCs are capable of self-renewal and differentiation into all types of embryonic and adult cells, including the germ-cell lineage. Therefore, gene targeting in mESCs enables reverse-genetic studies through the development of constitutive or conditional (using the Cre/LoxP system) KO mice. However, the classical way to isolate mouse ES cells is very inefficient. Several major improvements have greatly increased the success rate for deriving mESC lines, including the use of a defined serum-replacement (SR) medium15, alternating between mESC medium containing SR and fetal bovine serum (FBS)16, and the use of pharmacological compounds such as pluripotin or 2i17. Pluripotin, a small synthetic molecule, allows for the propagation of mESCs in an undifferentiated state in the absence of leukemia inhibitory factor (LIF) and mouse embryonic fibroblasts (MEFs)18. Finally, it has been shown that mESCs can be isolated with a very high efficiency (close to 100%) when an SR/FBS medium alternation protocol is combined with LIF and pluripotin19,20. These protocols enable the efficient isolation of RMCE-compatible KO mESCs that can subsequently be used for structure-function studies.
This paper describes a method that enables one to identify the key domains or residues within a protein that are responsible for specific cellular processes. To this end, a pipeline of advanced technologies that enable efficient mESC isolation, targeting vector assembly, and mESC targeting was created. As such, large panels with protein isoforms, domain mutants, and downstream effectors can be introduced in KO mESCs and can be evaluated for their ability to rescue the in vitro KO phenotype.
