Det er anslått at pattedyr-genomer inneholder omtrent 20.000 protein-kodende gener. Alternativ spleising og posttranslasjonelle modifikasjoner ytterligere øke protein repertoaret. Proteiner har en modulær konstruksjon 1 og ofte inneholde flere interaksjons domener, noe som tillater deres rekruttering i forskjellige proteinkomplekser og deres deltagelse i flere cellulære prosesser 2. Et eksempel er den multifunksjonelle proteiner kjent p120ctn. p120ctn kodes for av Ctnnd1 gen og består av et stort sentralt armadillo gjenta domene flankert av en N-terminal og en C-terminal region. Armadillo domene av p120ctn binder seg til en høyt konservert juxtamembrane domene av klassiske cadheriner som er involvert i celle-celle-adhesjon, men det binder også til den transkripsjon repressoren Kaiso. Det N-terminale domenet av p120ctn samvirker med forskjellige kinaser, fosfataser, små RhoGTPases og mikrotubul-assosiert proteins tre. Det er interessant som et resultat av alternativ spleising, p120ctn isoformer kan genereres fra fire alternative startkodoner 4. p120ctn isoform 1A er den lengste, som det er oversatt fra den mest-5' startkodon og inneholder full-lengde N-terminal segmentet. I p120ctn isoformer 3 og 4 er denne N-terminale segment slettes helt henholdsvis delvis og. Å forstå den nøyaktige rolle av proteiner (eller protein isoformene) og deres domenene i forskjellige cellulære funksjoner er fortsatt en utfordring.
Gen-målsøking i mESCs gjør det mulig å studere funksjonen til et protein gjennom den genetiske delesjon av det tilsvarende genet og har bidratt mye til identifisering av utviklingsmessig viktige og sykdom-relevante gener og veier. Dette gjennombrudd i revers genetikk var et resultat av fremskrittene innen feltene Mesc isolasjon og gen-målsøking som følge av homolog rekombinasjon 5 </sopp>. Homolog rekombinasjon er en prosess i hvilken DNA-fragmenter blir utvekslet mellom to lignende eller identiske nukleinsyrer grupper etter dobbelt-trådet (ds) DNA-brudd. Normalt er HR ineffektiv fordi dsDNA pauser er sjeldne. Nylig, kan effektiviteten av homologi-rettet gen-målsøking økes ved å bruke stedsspesifikke nuklease 6, 7, men dessverre disse er tilbøyelige til off-target effekter 8. En mer pålitelig teknikk for å muliggjøre gen-målsøking er RMCE, som er basert på setespesifikke rekombinasjon systemer som Cre / loxP eller FLPe / Frt. LoxP og Frt sekvens er funnet i bakteriofag P1 og Saccharomyces cerevisiae, henholdsvis, og består av 34 bp, inkludert et asymmetrisk 8 bp-sekvens som bestemmer orienteringen av området. På den annen side, kan orienteringen av, for eksempel, to loxP-seter innenfor et DNA strekning vil avgjøre hvorvidt den floxed DNA blir kuttet ut eller jegnversed ved Cre-formidlet rekombinasjon 9. Videre kan Cre også indusere en trans hvis to stedene ligger på forskjellige kromosomer. RMCE utnyttet heterospecific rekombinasjonsseter som ikke kryssreagerer og som er innebygd i et genom-locus. I nærvær av en donor plasmid som inneholder et DNA-fragment flankert av de samme heterospecific steder, vil den rekombinasen sette inn dette DNA-fragment i RMCE-kompatible genomiske lokuset på grunn av dobbel-samtidig translokasjon (figur 1). Her kan bare korrekt RMCE rettede kloner gjengi medikamentresistens, takket være en promoter på den innkommende vektoren som gjenoppretter en "fanget" promoter-mindre neomycin resistensgenet (neo R) som er tilstede i R26 genomet av dokkings-celler (Figur 1) 10, 11. Dette resulterer i en meget høy effektivitet målretting, ofte nær 100% 11 </ sup> 12. Som konklusjon, er RMCE basert målretting svært effektiv og kan anvendes for struktur-funksjoner studier; Men det krever en pre-utviklet genomisk locus.
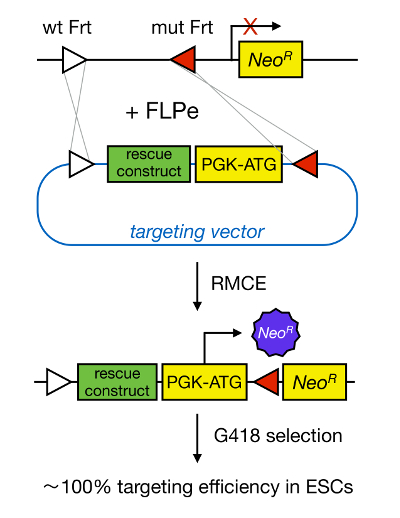
Figur 1. Skjematisk representasjon av RMCE-mediert målretting. RMCE tillater utveksling av DNA-segmenter fra en innkommende målsøkende vektor til en definert genomisk locus hvis begge huse to heterospecific FRT steder (vist med hvite og røde trekanter). I tillegg inneholder det konstruerte genomiske lokuset en promoterfrie og avkortet neomycin-resistens (Neo R) genet. Ved å tilveiebringe en promoter og startkodon i den innkommende DNA-fragment, kun korrekt rekombinasjonshendelser gjenopprette neomycinresistens, som resulterer i høye målretting effektivitet. Klikk her for å se en større versjon av thans skikkelse.
Genome engineering i mESCs tillater generering av RMCE-kompatibel mus. I 1981 ble to grupper lyktes å pluripotente celler fra den indre cellemasse (ICM) av blastocyster, og i å opprettholde dem i kultur 13, 14. mESCs er i stand til selvfornyelse og differensiering i alle typer av embryonale og voksne celler, inkludert bakterie-cellelinjen. Derfor, gen-målsøking i mESCs muliggjør revers-genetiske studier gjennom utviklingen av konstitutive eller betingede (ved bruk av Cre / loxP-system) KO-mus. Imidlertid er den klassiske metoden for å isolere muse-ES-celler er svært ineffektiv. Flere store forbedringer har sterkt øket suksessraten for utledning av Mesc linjer, inkludert bruk av et definert serum erstatning (SR) medium 15, alternerende mellom Mesc medium inneholdende SR og føtalt bovint serum (FBS) 16, og bruken av farmakologiskelogiske forbindelser slik som pluripotin eller 2i 17. Pluripotin, en liten syntetisk molekyl, tillater forplantning av mESCs i en udifferensiert tilstand i fravær av leukemi-inhiberende faktor (LIF) og muse-embryonale fibroblaster (MEFs) 18. Endelig har det vist seg at mESCs kan isoleres med en meget høy virkningsgrad (tilnærmet 100%) ved en SR / FBS-medium alternering protokoll er kombinert med LIF og pluripotin 19, 20. Disse protokollene muliggjøre effektiv isolering av RMCE-kompatible KO mESCs som senere kan brukes for struktur-funksjonsstudier.
Dette dokumentet beskriver en metode som gjør det mulig å identifisere de viktigste domener eller ester i et protein som er ansvarlig for spesifikke cellulære prosesser. For dette formål er en rørledning av avansert teknologi som muliggjør effektiv Mesc isolasjon, målsøkende vektor montasje, og Mesc målretting var skaped. Som sådanne store paneler med proteinisoformer, domene mutanter, og nedstrøms effektorer kan innføres i KO mESCs og kan evalueres for deres evne til å redde in vitro KO fenotype.
