In the present study we performed six ad-MVF isolation procedures with fat tissue from 7- to 12-month-old male wild-type C57BL/6 mice (mean body weight: 35 ± 1 g). Figure 1 illustrates the harvesting of murine epididymal fat pads with subsequent mechanical and enzymatic ad-MVF isolation. The time required for the harvesting of fat was 30 min and for the isolation of ad-MVF was 120 min. In total, the procedure took 150 min.
We harvested 1.2 ± 0.1 mL of adipose tissue per donor animal. This amount of fat allowed the isolation of 42,000 ± 2,000 ad-MVF per mL. The mean length of the isolated ad-MVF was 42 ± 1 µm (Figure 2A). The duration of enzymatic digestion was determined by means of microscopic control as shown in Figure 2B. ad-MVF exhibited a typical microvessel morphology with hierarchical vessel segments (Figure 2C).
We additionally characterized the ad-MVF by means of flow cytometry. For this purpose, ad-MVF were further digested in cell detachment solution for 30 min into single cells15. The flow cytometric analyses revealed that ad-MVF contain 26 ± 2% CD31-positive endothelial cells, 17 ± 2% α-smooth muscle actin (SMA)-positive perivascular cells, and 9 ± 1% cells positive for the mesenchymal stem cell marker CD117 (Figure 3A-C). After seeding on a dermal skin substitute, which was fixed and sectioned immediately for histological analyses, larger ad-MVF were mainly localized on the implant's surface (Figure 3D). However, capillary vessel segments could also be detected within its pores. Immunohistochemical staining of seeded ad-MVF further revealed physiological microvessel configuration with arteriolar, venular and capillary-like fragments (Figure 3E).
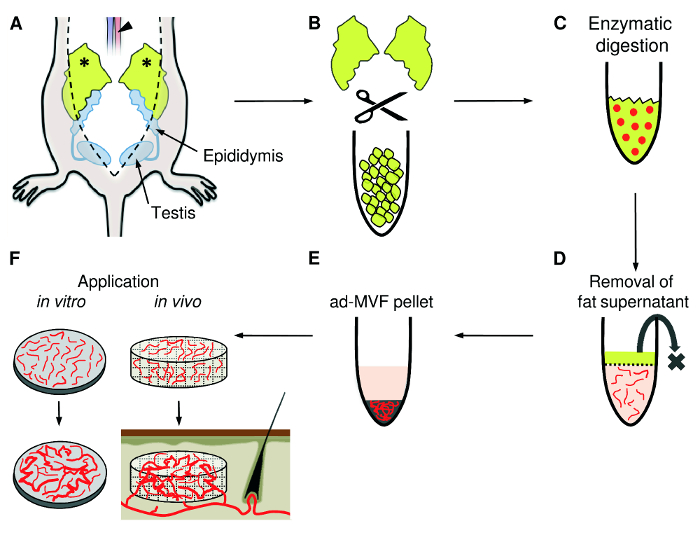
Figure 1: ad-MVF Isolation. A) Following midline laparotomy, the epididymal fat pads (asterisks) are identified. The adipose tissue is harvested with a small preparation scissors and the animal is sacrificed by incision of the abdominal aorta (arrowhead). B) Mechanical mincing of the fat pads with fine scissors until the tissue suspension appears homogeneous. C) For enzymatic digestion, collagenase is added to the tissue suspension. D) The cell-vessel suspension is incubated and the supernatant fat layer is discarded. E) After centrifugation, the pellet containing ad-MVF is resuspended and can be used for different applications, such as the in vitro analysis of microvascular network formation or the seeding of ad-MVF on scaffolds for in vivo implantation into tissue defects (F). Please click here to view a larger version of this figure.
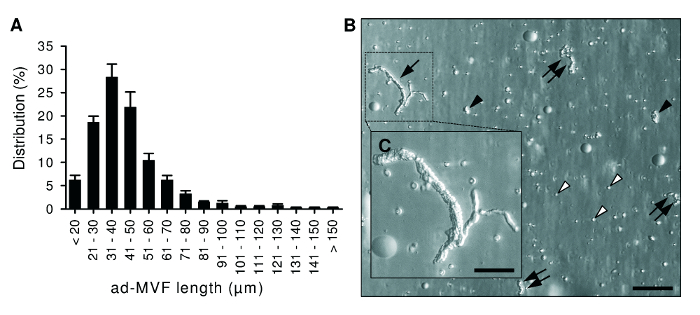
Figure 2: Morphological Characteristics of ad-MVF. A) Length distribution of freshly isolated ad-MVF. Mean ± standard error of the mean (n = 6). B) Microscopic image of a cell-vessel suspension smear with large (arrow), medium-sized (double arrows) and small (black arrowheads) ad-MVF as well as single cells (white arrowheads). Scale bar = 110 µm. Higher magnification reveals that ad-MVF exhibit a mature microvessel morphology with hierarchical microvessel segments (C, inset of B). Scale bar: C = 50 µm. Please click here to view a larger version of this figure.

Figure 3: Flow Cytometric, Histological and Immunohistochemical Characteristics of ad-MVF. A-C) Flow cytometric analyses of freshly isolated ad-MVF illustrating CD31-positive endothelial cell (A), α-SMA-positive perivascular cell (B) and CD117-positive mesenchymal stem cell (C) fractions. Appropriate isotype-identical controls were used to adjust threshold levels. Mean ± standard error of the mean (n = 6). D) Hematoxylin and eosin-stained section of a seeded dermal skin substitute. A few capillary vessel segments are located in the implant's pores (arrowheads). Larger microvessels with venular (double arrow) or arteriolar (arrow) morphology are localized on its surface. Scale bar = 40 µm. Broken line = implant border. E) Characterization of ad-MVF by means of immunohistochemical detection of the endothelial cell marker CD31 (red) and the perivascular cell marker α-SMA (green). Cell nuclei were stained with a DNA-binding fluorescent dye (blue). Arteriolar ad-MVF (arrow) are characterized by a thicker α-SMA-positive cell layer when compared to venular fragments with a larger lumen (double arrow). Arrowheads = capillary ad-MVF. Scale bar = 25 µm. Please click here to view a larger version of this figure.
