Sistema nervoso central (SNC) tem uma capacidade limitada para compensar a perda e/ou disfunção dos neurônios e vias axonal que acompanham as condições tais como traumatismo crânio-encefálico (TCE), derrame, medular lesão (SCI) e neurodegenerativas doença1 ,2,3,4,5. Neurogênese no SNC é restrito a um número limitado de áreas no cérebro, impedindo a restauração de neurônios perdidos6,7. Além disso, a regeneração das vias axonal perdidas no SNC é insuficiente devido a falta de orientação dirigida, a presença de inibidores de consequência natural e astrogliosis reativa após danos ao tecido neural2,8, 9,10. Astrócitos normalmente têm diversas funções na assistência neurônios com homeostase iónica, liberação de neurotransmissores, formação de sinapse e neurovasculares acoplamento11. No entanto, seguindo até leve dano ao tecido neural, astrócitos podem sofrer mudanças moleculares, estruturais e funcionais como eles transição para um estado hipertrófica11. Em resposta à severa neurotrauma, estas mudanças resultam na formação de uma cicatriz com uma penumbra contendo migrando astrócitos reativos e um núcleo de lesão que inclui leucócitos vazados da ruptura barreira hemato – encefálica (BBB), microglia, oligodendrócitos e fibroblastos11,12,13. Estes astrócitos reativos atingir uma morfologia de processos filamentosos, desorganizados e exibem expressão aumentada de proteínas de filamento intermediário e proteoglicanos de sulfato de condroitina (CSPGs), que impedem a regeneração neural12. Mesmo que a cicatriz glial inicialmente ajuda a restaurar a integridade BBB e evitar a transmissão da resposta inflamatória ao tecido saudável circundante, serve como uma barreira física e bioquímica contra axônio regeneração12,14 ,15,16. Por exemplo, axônios que encontrar a cicatriz glial exibir cones de crescimento distrófica bulbosa e atrofiado crescimento12. Além disso, a desorganização dos processos hamartomas após lesão impede a extensão de regeneração de axônios17. O resultado destas características inibitório manifesta-se nas deficiências permanentes muitas vezes físicas e neurológicas que os pacientes sofrem após grave neurotrauma, incluindo TBI e Sci.
Independentemente dos extrínsecos desafios funcional regeneração no SNC, axônios demonstraram possuir uma capacidade intrínseca de se regenerar. Por exemplo, a natureza dinâmica dos cones distrófica crescimento em contato com a cicatriz glial sugere que estes finais mantém sua capacidade de estender12. Por conseguinte, acredita-se que um obstáculo principal para a re-crescimento axonal é o ambiente inibitório do CNS pós-lesão e que fornecendo um ambiente mais permissivo via redução de cicatrizes e/ou fornecendo regenerativas pontes sobre a cicatriz seria gliais vantajoso. De fato, estudos anteriores demonstraram que os neurônios do CNS eram capazes de estender axônios através de uma lesão utilizando enxertos de nervo periférico como pontes, que apresentam um ambiente mais favorável para o axônio regeneração12,18, 19. Seguiram-se diversas outras estratégias para explorar essa capacidade regenerativa vestigial. Por exemplo, manipulação de vias de sinalização celular crescimento em vários modelos de lesão resultou na regeneração axonal e cicatriz glial redução10,20,21. Além disso, estudos têm mostrado que o tratamento com chondroitinase ABC, que fende a maioria das cadeias de açúcar em CSPGs, diminui o efeito inibitório de CSPGs secretada por astrócitos reativos22. Apesar de encorajador resultados, essas abordagens não fornecem dirigido a orientação dos cones de crescimento, que pode potencialmente resultar em regeneração aberrante12e também não conta para a perda de neurônios. Abordagens baseadas em célula têm sido utilizadas em tentativas de ultrapassar os efeitos da cicatriz glial e para repor as células perdidas, particularmente os neurônios. Alguns grupos têm os astrócitos reativos em neurônios, enquanto outros transplantamos células progenitoras neurais em lesões de CNS para repovoar a área de lesão e promover o axônio regeneração23,24, 25. no entanto, transplante de células-tronco sozinho é limitado por taxas de sobrevivência baixa integração pobre e modesta de retenção no tecido danificado5. Além disso, estas estratégias baseadas em células não restaurar a longa distância axonal panfletos, especialmente em uma maneira controlada. Portanto, biomateriais em combinação com outras abordagens estão sendo exploradas como veículos de entrega para vários neural e fatores de crescimento e células progenitoras26. Abordagens baseadas em biomaterial apresentam um alto grau de controle de projeto para produzir construções que imitam o haptotaxic físico, específico, e sinais quimiotáxicas presente no microambiente (3D) tridimensional do destino anfitrião tecido27, 28,29,30,31,32,33,34. Reprodução desses sinais ambientais é de suma importância para células transplantadas apresentar morfologia como de um nativo, proliferação, migração e sinalização, entre outras características neurobiológicas29. Apesar dessas propriedades vantajosas, avanço além da tradicional célula semeada biomaterial andaimes é necessário para substituir neurônios perdidos e promover a regeneração axonal longa distância dirigida simultaneamente.
Uma promissora abordagem alternativa baseia-se no tecido neural engenharia “andaimes vivos”, que são distintos de outras abordagens baseadas em célula devido à presença de células neurais vivas com um cytoarchitecture pré-formadas que emula neuroanatomia nativa e/ou mecanismos do desenvolvimento para facilitar a substituição do alvo, reconstrução e regeneração dos circuitos neurais4,35. Considerações para o projeto de vida andaimes incluem os fenótipos e fontes de células neurais, bem como as propriedades mecânicas e/ou físicas e os sinais bioquímicos ditaram pela composição de qualquer acompanhamento de biomateriais35. Após a fabricação em vitro, estes andaimes vivos podem ser implantado no vivo de moléculas de adesão celular presente e Quimiotáticos e neurotrophic sinaliza para regular ativamente a migração de células nervosas e consequência natural do axônio dependendo do estado e progressão dos processos regenerativos35. Células gliais podem servir como base para a cytoarchitecture engenharia de andaimes de vida, desde que estas células mediam diversos mecanismos do desenvolvimento na vivo. Durante o desenvolvimento do cérebro, os neurônios novos dependem de processos basais prorrogados pela glia radial da zona ventricular para a placa de desenvolvimento cortical como andaimes vivos para migração dirigida36,37. Além disso, estendendo o crescimento cones são mostrada para orientar-se por sensoriamento de sinais atraentes e repelentes eliciados células gliais guidepost, and so-called “pioneiro” axônios são sugeridos para alcançar as metas corretas, estendendo-se ao longo de pre-modelado gliais moldes de38,35,39. Assim, as células gliais são necessárias para a orientação do pioneirismo de axônios, que mais tarde servem como baseado no axônio “andaimes de vida” para direcionar a projeção de axônios “seguidor”. Além disso, mecanismos de crescimento mediada por glia foram mostrados para persistir pós-natal, como neuroblastos seguem o fluxo migratório rostral (RMS) para navegar na zona subventricular (SVZ), uma das poucas áreas remanescentes da neurogênese no cérebro adulto, para o bulbo olfatório (OB)40. Estes neuroblastos em RMS migram dentro do tubo glia (figura 1A-1), que é composto por processos hamartomas alinhados longitudinalmente, através de aderências direta célula-célula e localizadas fatores solúveis37, 41. por fim, enquanto danos CNS em causas de mamíferos interrompeu arranjo hamartomas processo formando uma cicatriz glial que impede fisicamente a regeneração axonal17, muitos sistemas não-mamíferos faltam a formação de uma cicatriz glial prejudicial. Em vez disso, as células gliais de espécies não-mamíferos mantenham mais organizado, alinhados a padrões que são usados como guias através da região lesada17,42,43. Por exemplo, em modelos de não-mamíferos SCI, axônios são mostrados para crescer em estreita associação com gliais pontes cruzando a lesão, sugerindo um papel importante para andaimes gliais organizados como substratos, facilitando a regeneração axonal e recuperação funcional ( Figura 1A -2) 42 , 44 , 45. recapitulação das características neuroanatômica e os mecanismos do desenvolvimento/regenerativa descritos acima pode produzir uma nova classe de engenharia baseada em gliais vivos andaimes que simultaneamente pode conduzir a migração neuronal imatura e axonal Pathfinding através de ambientes caso contrário não-permissivo, assim, potencialmente, mitigação dos efeitos da neuronal e degeneração do trato de axônio associado com lesão do CNS e doença.
Nosso grupo de pesquisa anteriormente desenvolveu vários tipos de andaimes de vida para a reconstrução e regeneração dos tracts axonal no SNC e o sistema nervoso periférico (SNP) através de microtecido projetado redes neurais (micro-TNS) e tecido enxertos de nervo engenharia (TENGs), respectivamente,27,46,,47,48. Ambas as estratégias são inerentemente baseia biomimetismo. Micro-TNS são estruturas anatomicamente inspirada projetadas estruturalmente e funcionalmente substituir axonal folhetos conectando distintas populações neuronais do cérebro. TENGs explorar o mecanismo de desenvolvimento da regeneração axonal axônio-facilitada, exemplificado pelo crescimento do axônio “seguidor” ao longo de axônios “pioneiro”, para alcançar o hospedeiro alvo regeneração axonal35,46,48. Nós recentemente capitalizou a versatilidade do andaime a vida técnica usando um esquema similar do encastoamento como micro-TNS e buscando inspiração os mecanismos baseados em glia de presentes durante todo o desenvolvimento. Aqui, nós desenvolvemos construções consistindo de feixes hamartomas alinhados, abrangendo o lúmen colágenas de um hidrogel microcoluna49. Estes andaimes hamartomas vivos são desenvolvidos pelo primeiro preencher um conjunto de agulha de acupuntura-tubo capilar com líquido agarose para criar um hidrogel cilíndrico oco com um diâmetro externo (OD) e o diâmetro interno (ID) correspondente para os diâmetros do tubo e agulha, respectivamente. Na sequência de gelificação de agarose e extração de hidrogel microcoluna do tubo capilar, o interior oco é revestido com tipo eu colágeno para fornecer um ambiente permissivo para a adesão de astrocyte e alinhado à formação de feixe (figura 1B -1). Depois, o lúmen é semeado com astrócitos corticais cerebrais, isolados de filhotes pós-natal (figura 1B-2). Contrariamente ao bidimensional (2D) alinhamento as técnicas que dependem da aplicação de campos elétricos, micropatterned grooves e matriz extracelular da proteína (ECM), padronização, alinhamento astrocyte no cadafalso vivos técnica depende de auto-montagem de acordo com variáveis controláveis como colágeno concentração50,,51,52, densidade celular e curvatura do substrato (coluna ID). Os astrócitos contraem e remodelam o colágeno e adquirem uma morfologia bipolar, alinhado longitudinalmente análoga para os andaimes naturais observados na vivo (figura 1B-3). Com efeito, estamos ativamente perseguindo o uso destas estruturas de cabo, como como substratos físicos para orientação alvo da migração de neurônios imaturos, bem como facilitar a regeneração axonal através do ambiente desfavorável do SNC danificado, particularmente a cicatriz glial mamíferos (Figura 1). Este artigo irá apresentar o método de fabricação detalhada para as microcolunas hamartomas, contraste e imunofluorescência imagens de cytoarchitecture o esperado e uma discussão abrangente sobre as limitações atuais e futuras direções da fase a técnica.
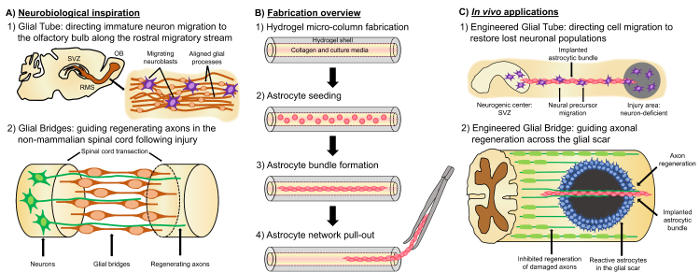
Figura 1: inspiração, protocolo de fabricação e aplicações propostas para as redes de hamartomas alinhadas. (A) neurobiológicos inspiração: neuroblastos (1), originários da zona subventricular neurogênica (SVZ) utilizam o tubo glia alinhado longitudinalmente do fluxo migratório rostral (RMS) para migração dirigida para o bulbo olfatório (OB); (2) não-mamíferos, como os anfíbios e peixes podem sustentar a regeneração após dano no tecido neural em parte devido à formação de uma ponte glia que conecta as extremidades de uma lesão (por exemplo, medula espinhal necrosante) e serve como um andaime para a orientação de regeneração de axônios. (B) visão geral de fabricação: (1) a construção de uma hidrogel de micro-empresas, oco microcoluna com o lúmen revestido com ECM, (2) semeadura do primários astrócitos corticais isolados de filhotes pós-natal, (3) auto-montagem do longitudinalmente orientada pacotes em cultura e (4) extraído do pacote o invólucro de biomaterial para estudos futuros de implantação. (C) na vivo aplicações: (1) estes andaimes de vida podem servir como tubos gliais projetados para a migração de neurônio dirigido de centros neurogênicos repovoar regiões do neurônio-deficiente; (2) recapitulação o mecanismo do desenvolvimento de pioneirismo orientação do axônio e o mecanismo regenerativo das pontes gliais de não-mamíferos pode dotar estes andaimes hamartomas com a capacidade de direcionar a regeneração do axônio através do não-permissivo ambiente da cicatriz glial mamífera. Clique aqui para ver uma versão maior desta figura.
