Das zentrale Nervensystem (ZNS) hat eine begrenzte Kapazität gegen Verlust und/oder Funktionsstörungen der Nervenzellen und axonalen Signalwege, die Bedingungen wie Schädel-Hirn-Verletzungen (SHT), begleiten Schlaganfall, Rückenmark Verletzungen (SCI) und Neurodegenerative Krankheit1 ,2,3,4,5. Neurogenese im ZNS beschränkt sich auf eine begrenzte Anzahl von Bereichen im Gehirn behindern die Wiederherstellung der verlorenen Neuronen6,7. Darüber hinaus ist Regeneration der verlorenen axonalen Wege im ZNS nicht ausreichend durch das Fehlen einer gezielten Führung, das Vorhandensein von Auswuchs Inhibitoren und reaktive Astrogliosis nach Schäden an Nervengewebe2,8, 9,10. Astrozyten haben in der Regel diverse Funktionen bei der Unterstützung der Neuronen mit Ionen-Homöostase, Neurotransmitter-Clearance, Synapse Bildung und neurovaskuläre Kopplung11. Dennoch können Astrozyten nach auch leichte Schäden an Nervengewebe, molekulare, strukturelle und funktionelle Veränderungen unterziehen, um eine hypertrophe staatliche11Übergang. Als Reaktion auf schwere Neurotrauma führen diese Veränderungen bei der Bildung einer Narbe mit einer Penumbra, Migration von reaktiven Astrozyten und eine Läsion Kern, der Leukozyten von der geplatzten Blut – Hirn-Schranke (BBB), Mikroglia durchgesickert enthält enthält, Oligodendrozyten und Fibroblasten11,12,13. Diese reaktive Astrocyten erreichen eine Morphologie des faserigen, ungeordnete Prozesse und zeigen erhöhte Expression von intermediate Filament Proteine und Chondroitinsulfat Proteoglycans (CSPGs), die neurale Regeneration12behindern. Obwohl die glial Narbe anfangs hilft BBB Integrität wiederherzustellen und Übertragung der Entzündungsreaktion umliegenden gesunden Gewebes zu vermeiden, dient es als biochemische und physikalische Barriere gegen Axon Regeneration12,14 ,15,16. Zum Beispiel Axone, die Begegnung die glial Narbe bauchige dystrophischen Wachstum Kegel anzeigen und verkümmert Wachstum12. Darüber hinaus behindert die Desorganisation der astrocytic Prozesse nach einer Verletzung die Verlängerung der regenerierende Axone17. Das Ergebnis dieser hemmenden Eigenschaften manifestiert sich in der oft dauerhafte körperlichen und neurologischen Beeinträchtigungen, die Patienten nach schweren Neurotrauma leiden, einschließlich TBI und Sci.
Unabhängig von der äußeren Herausforderungen funktionelle Regeneration im ZNS nachweislich die Axone besitzen eine intrinsische Fähigkeit zu regenerieren. Zum Beispiel zufolge die dynamische Natur der dystrophischen Wachstum Kegel in Kontakt mit der glial Narbe diese Endungen behalten ihre Fähigkeit,12zu erweitern. Infolgedessen wird es vermutet, dass als wichtigste Hindernis für axonalen Nachwachsen der hemmenden Umwelt der posttraumatischen CNS und Bereitstellung einer großzügigeren Umgebung über Reduktion glialen Narbenbildung und/oder sofern regenerative Brücken über die Narbe wäre vorteilhaft. In der Tat haben Studien gezeigt, dass ZNS-Neuronen waren in der Lage, die Ausweitung der Axone durch eine Läsion mit peripheren Nerv Transplantationen als Brücken, die ein günstigeres Umfeld für Axon Regeneration12,18zu präsentieren, 19. Mehrere andere Strategien haben verfolgt, um diese verkümmerte Regenerationsfähigkeit zu nutzen. Manipulation der Zelle Wachstum Signalwege in verschiedenen Verletzungen resultieren beispielsweise axonalen Regeneration und glial Narbe Reduktion10,20,21. Darüber hinaus haben Studien gezeigt, dass die Behandlung mit Chrondroitinase ABC, die den Großteil der Zuckerketten in CSPGs zerspaltet, die hemmende Wirkung von CSPGs abgesondert durch reaktive Astrocyten22verringert. Trotz ermutigender Ergebnisse, diese Ansätze bieten keine Führung des Wachstums Zapfen, gerichtet, die potenziell aberrierende Regeneration12führen kann, und auch nicht für den Verlust von Neuronen zu berücksichtigen. Zell-basierte Ansätze wurden genutzt, bei versuchen, die Auswirkungen der glial Narbe zu überwinden und um verlorene Zellen, vor allem Neuronen wieder aufzufüllen. Einige Gruppen haben entdifferenzierten reaktive Astrocyten in Neuronen, während andere neurale Vorläuferzellen in CNS Läsionen zum repopulate Bereich Verletzungen und zur Förderung Axon Regeneration23,24, , verpflanzt haben 25. Stammzelltransplantation allein ist jedoch durch die niedrigen Überlebensraten, mangelhafte Integration und bescheidenen Retention in das geschädigte Gewebe5begrenzt. Darüber hinaus nicht diese Zell-basierte Strategien Fernverkehr axonalen Landstriche, vor allem in einer kontrollierten Weise wiederherzustellen. Daher als Lieferfahrzeuge für verschiedene neuronale Biomaterialien in Kombination mit anderen Ansätzen erforscht und Vorläuferzellen und Wachstum Faktoren26. Biomaterial-basierte Ansätze verfügen über ein hohes Maß an Design-Steuerung, Konstrukte zu produzieren, die die spezifischen körperlichen, Haptotaxic zu imitieren, und chemotaxic Hinweise in der dreidimensionalen (3D) Mikroumgebung der Ziel-Host Gewebe27, 28,29,30,31,32,33,34. Reproduktion von diesen Umweltsignale ist von größter Bedeutung für die transplantierten Zellen, Native-ähnliche Morphologie, Proliferation, Migration und Signaltechnik, unter anderen neurobiologischen Eigenschaften29zu präsentieren. Trotz dieser vorteilhaften Eigenschaften ist Fortschritt jenseits traditioneller Zelle ausgesät Biomaterial Gerüste erforderlich, um gleichzeitig Regie Fernverkehr axonalen Regeneration fördern und ersetzen verlorene Neuronen.
Eine vielversprechende alternative Ansatz basiert auf Nervengewebe entwickelt “lebenden Gerüste”, unterscheidet sich von anderen Zell-basierte Ansätze aufgrund des Vorhandenseins von lebenden Nervenzellen mit einem vorgeformten Cytoarchitecture, die native Neuroanatomie emuliert und/oder Entwicklungsmechanismen gezielte Ersatz, Wiederaufbau und Regenerierung der neuralen Schaltkreis4,35zu erleichtern. Überlegungen für die Gestaltung von lebendigen Gerüste gehören die Phänotypen und Quellen von neuronalen Zellen sowie die mechanische/physikalische Eigenschaften und die biochemische Signale durch die Zusammensetzung der begleitenden Biomaterialien35diktiert. Nach Fertigung in Vitro, diese lebenden Gerüste können implantiert in Vivo vorhanden Zelle Adhäsionsmoleküle und chemotaktische und neurotrophe Signale zum neuronalen Zellwanderung und Axon Auswuchs je nach Zustand aktiv regulieren und Fortschreiten der regenerativen Prozesse35. Gliazellen dienen als Grundlage für die veränderter Cytoarchitecture lebenden Gerüste, da diese Zellen verschiedene Entwicklungsmechanismen in Vivozu vermitteln. Während der Entwicklung des Gehirns setzen neue Neuronen auf basalen Prozesse durch radiale Gliazellen aus der ventrikulären Zone in Richtung der Entwicklung kortikale Platte als lebende Gerüste für gerichtete Migration36,37erweitert. Darüber hinaus erweitern Wachstum sind Zapfen gezeigt, sich zu orientieren, durch Sensierung attraktiv und abweisend Signale ausgelöst durch Wegweiser Glia-Zellen, und sogenannte “Pionierarbeit” Axone werden vorgeschlagen, um die richtigen Ziele zu erreichen, durch die Ausdehnung entlang Pre-gemusterten Glia 35,38,39Gerüste. Gliazellen sind also notwendig, um die Führung des bahnbrechenden Axone, die später als Axon-basierte dienen “lebenden Gerüste” um die Projektion der “Mitläufer” Axone leiten. Darüber hinaus Glia-vermittelten Wachstum Mechanismen haben gezeigt, dass postnatal, fortbestehen folgendermaßen Neuroblasten rostral wandernden Stream (RMS), aus der subventricular Zone (SVZ), einer der wenigen verbliebenen Bereiche der Neurogenese im erwachsenen Gehirn zu navigieren die Riechkolben (OB)40. Diese Neuroblasten in der RMS migrieren innerhalb der Glia Röhre (Abbildung 1A-1), die besteht aus längs ausgerichteten astrocytic Prozesse, über direkten Zell-Zell-Verwachsungen und lokalisiert lösliche Faktoren37, 41. schließlich während CNS Schäden in Säugetieren Ursachen astrocytic Prozess Anordnung bilden eine glial Narbe, die körperlich axonale Regeneration17 gestört behindert, viele nicht-Säugetier-Systemen fehlt die Bildung einer nachteiligen glial Narbe. Vielmehr pflegen Gliazellen des nicht-Säugetier-Arten mehr organisiert, ausgerichtet von Mustern, die als Führer durch die verletzte Region17,42,43verwendet werden. Zum Beispiel nicht-Säugetier-SCI-Modelle, Axone entnehmen Sie bitte weiter in enger Zusammenarbeit mit Glia Brücken über der Läsion, schlägt eine wichtige Rolle für organisierte Glia Gerüste als Erleichterung der axonalen Regeneration und funktionelle Wiederherstellung (Substrate Abbildung 1A -2) 42 , 44 , 45. Reprise der neuroanatomischen Eigenschaften und die Entwicklungs-/regenerative Mechanismen, die oben beschriebenen kann dies zu eine neue Klasse von veränderter Glia-basierte lebenden Gerüste, die unreif neuronalen Migration gleichzeitig fahren können und axonalen Wegfindung durch ansonsten freizügigen Umgebungen, damit potenziell Minderung der Auswirkungen von neuronalen und Axon Trakt Degeneration mit CNS Verletzung und Krankheit verbunden.
Unsere Forschungsgruppe hat bereits mehrere Arten von lebenden Gerüste für den Wiederaufbau entworfen und Regeneration der axonalen Traktate in das ZNS und das periphere Nervensystem (PNS) über Micro-Gewebe entwickelt, neuronale Netze (Mikro-TENNs) und Gewebe Nerv Transplantationen (TENGs) bzw.27,46,47,48entwickelt. Beide Strategien basieren grundsätzlich auf Biomimicry. Mikro-TENNs sind anatomisch inspirierte Strukturen strukturell und funktionell axonalen Traktate verbinden unterschiedliche neuronale Populationen des Gehirns ersetzen soll. TENGs ausnutzen den Entwicklungsbiologie Mechanismus der Axon erleichtert axonalen Regeneration, veranschaulicht durch “Mitläufer” Axon Wachstum entlang “Pionier” Axone, gezielte Host axonalen Regeneration35,46,48zu erreichen. Wir vor kurzem auf die Vielseitigkeit des Gerüstes lebenden aktiviert Technik mit einem ähnlichen Schema Ummantelung als Mikro-TENNs und inspirieren von den Glia-basierte Mechanismen zu präsentieren, während der gesamten Entwicklung. Hier haben wir Konstrukte bestehend aus ausgerichteten astrocytic Bündel überspannt das kollagene Lumen ein Hydrogel Mikro-Spalte49entwickelt. Diese astrocytic lebenden Gerüste werden von ersten füllen eine Kapillare Rohr-Akupunktur Nadel Montage mit flüssige Agarose Erstellen einer hohlen zylindrischen Hydrogel mit einem Außendurchmesser (OD) und Innendurchmesser (ID) entspricht der Durchmesser der entwickelt die Schlauch und Nadel, beziehungsweise. Nach Agarose Gelierung und Extraktion der Hydrogel Mikro-Spalte aus der Kapillare, die innen hohl ist beschichtet mit Typ ich Kollagen, ein Umfeld für Astrozyten Adhäsion permissive liefern und ausgerichtet Bundle Bildung (Abbildung 1 b -1). Danach ist das Lumen mit zerebrale kortikale Astrozyten isoliert vom postnatalen Ratte Welpen (Abbildung 1 b-2) ausgesät. Im Gegensatz zu zweidimensionalen (2D) Ausrichtung Techniken, die auf die Anwendung von elektrischen Feldern, Micropatterned Rillen und extrazelluläre Matrix (ECM) Protein-Strukturierung, Astrozyten Ausrichtung in die lebendige Gerüst stützt sich Technik auf Selbstmontage nach steuerbare Variablen wie Substrat Krümmung (Spalte ID), Zelldichte und Kollagen Konzentration50,51,52. Die Astrozyten Vertrag umzugestalten das Kollagen und erwerben eine bipolare, längs ausgerichtet Morphologie, die analog zu den natürlichen Gerüste in Vivo (Abbildung 1 b-3) beobachtet. In der Tat verfolgen wir aktiv den Einsatz dieser Kabel-ähnliche Strukturen als physische Substrate für gezielte Führung des Migration unreifer Nervenzellen sowie Erleichterung der axonalen Regeneration durch die ungünstigen Umfeld des beschädigten ZNS, insbesondere die Säugetier-glial Narbe (Abbildung 1). Dieser Artikel wird die detaillierte Herstellungsverfahren für die astrocytic Mikro-Spalten präsentieren, phase Kontrast und Immunfluoreszenz Bilder von der erwarteten Cytoarchitecture und eine umfassende Diskussion über die aktuellen Beschränkungen und zukünftige Richtungen von der Technik.
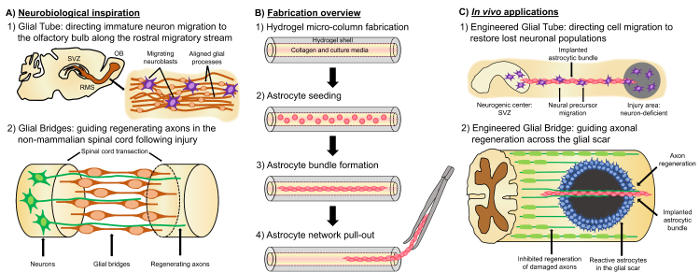
Abbildung 1: Inspiration, Herstellung Protokoll und vorgeschlagenen Anwendungen für die ausgerichteten Astrocytic Netzwerke. (A) neurobiologische Inspiration: (1) Neuroblasten aus der neurogenen subventricular Zone (SVZ) nutzen das längs ausgerichtete Glia Rohr in den rostral wandernden Stream (RMS) für gerichtete Migration in den Riechkolben (OB); (2) nicht-Säugetiere wie Amphibien und Fischen verträgt Regeneration nach Nervengewebe Schäden teilweise durch die Bildung einer Glia Brücke, die verbindet die Enden einer Läsion (z.B. durchtrennten Rückenmark) und dient als Gerüst für die Führung des regenerierende Axone. (B) Herstellung Übersicht: (1) Bau einer Mikrometer Größe, hohlen Hydrogel Mikro-Spalte mit dem Lumen mit ECM, beschichtet (2) Aussaat von primären kortikalen Astrozyten isoliert vom postnatalen Ratte Welpen (3) Selbstmontage der längs-orientierte Bündel in Kultur und (4) Extraktion des Bundles aus Biomaterial-Ummantelung für zukünftige Implantation Studien. (C) In Vivo Anwendungen: (1) diese lebenden Gerüste können dienen als veränderter Glia Rohre für gerichtete Neuron Migration von neurogenen Zentren Neuron-defizienten Regionen; neu aufzufüllen (2) Reprise des developmental Mechanismus von zukunftsweisenden Axon Guidance und der regenerativen Mechanismus der Glia Brücken in nicht-Säugetiere kann diese astrocytic Gerüste mit einer Kapazität von Axon Regeneration über die non-permissive direkte verleihen. Umgebung der Säugetier-glial Narbe. Bitte klicken Sie hier für eine größere Version dieser Figur.
