Il sistema nervoso centrale (SNC) ha una capacità limitata per contrastare la perdita e/o disfunzione dei neuroni e vie axonal che accompagnano condizioni come la ferita di cervello traumatica (TBI), ictus, malattia di ferita (SCI) e neurodegenerative del midollo spinale1 ,2,3,4,5. Neurogenesi nel SNC è limitata a un numero limitato di aree del cervello, che ostacolano il restauro di neuroni persi6,7. Inoltre, la rigenerazione delle vie axonal perse nello SNC è insufficiente a causa della mancanza di orientamento diretto, la presenza di inibitori di conseguenza e astrogliosi reattiva dopo danno di tessuto neurale2,8, 9,10. Astrociti in genere hanno diverse funzioni nell’assistere i neuroni con omeostasi ionica, neurotrasmettitore gioco, formazione della sinapsi e neurovascular attacco11. Tuttavia, in seguito anche lievi danni al tessuto neurale, astrocytes possono subire variazioni molecolari, strutturali e funzionali come la transizione a un stato ipertrofico11. In risposta a grave neurotrauma, questi cambiamenti provocano la formazione di una cicatrice con una penombra contenente la migrazione astrociti reattivi e un nucleo di lesione che include leucociti trapelati dalla rottura emato – encefalica (BBB), microglia, oligodendrociti e fibroblasti11,12,13. Questi astrociti reattivi raggiungono una morfologia dei processi filamentosi, disorganizzati ed esibiscono l’espressione aumentata di proteine dei filamenti intermedi e proteoglicani Condroitin solfato (CSPG), che ostacolano la rigenerazione neurale12. Anche se inizialmente la cicatrice gliale aiuta a ripristinare l’integrità BBB ed evitare la trasmissione della risposta infiammatoria al tessuto sano circostante, serve come una barriera fisica e biochimica contro assone rigenerazione12,14 ,15,16. Per esempio, gli assoni che incontrano la cicatrice gliale visualizzare dei coni di crescita distrofica bulboso e stentata crescita12. Inoltre, la disorganizzazione dei processi astrocitari dopo lesione impedisce l’estensione della rigenerazione di assoni17. Il risultato di queste caratteristiche inibitorie si manifesta spesso permanenti menomazioni fisiche e neurologiche che pazienti soffrono dopo neurotrauma grave, tra cui TBI e SCI.
Indipendentemente dalle sfide estrinseche funzionale rigenerazione nel SNC, assoni hanno dimostrati di possedere un’intrinseca capacità di rigenerarsi. Per esempio, la natura dinamica dei coni di crescita distrofico a contatto con la cicatrice gliale suggerisce che queste terminazioni mantengono la loro capacità di estendere12. Di conseguenza, si ritiene che un ostacolo principale alla ricrescita assonale è l’ambiente inibitorio del SNC di alberino-ferito e che fornendo un ambiente più permissivo via la riduzione di cicatrici e/o fornendo rigenerative ponti attraverso la cicatrice sarebbe glial vantaggioso. Infatti, studi precedenti hanno dimostrato che i neuroni CNS erano capaci di estendere gli assoni attraverso una lesione usando gli innesti di nervi periferici come ponti, che presentano un ambiente più favorevole per assone rigenerazione12,18, 19. Parecchie altre strategie sono state perseguite per sfruttare questa capacità rigenerativa vestigiale. Ad esempio, manipolazione delle vie di segnalazione cellulare crescita in vari modelli di lesione ha provocato la rigenerazione assonale e cicatrice gliale riduzione10,20,21. Inoltre, gli studi hanno indicato che il trattamento con chondroitinase ABC, che scinde la maggior parte delle catene dello zucchero in CSPG, diminuisce l’effetto inibitorio di CSPG secernuto da astrociti reattivi22. Nonostante incoraggianti i risultati, questi approcci non forniscono diretto orientamento dei coni di crescita, che può potenzialmente causare rigenerazione aberrante12e inoltre non tengono conto della perdita di neuroni. Approcci basati su cellule sono stati utilizzati nei tentativi per superare gli effetti della cicatrice gliale e per ricostituire le cellule perse, in particolare i neuroni. Alcuni gruppi hanno dedifferenziate di astrociti reattivi nei neuroni, mentre gli altri sono trapiantate cellule progenitrici neurali in lesioni dello SNC di ripopolare la zona ferita e promuovere assone rigenerazione23,24, 25. Tuttavia, trapianto di cellule staminali è limitato dalla bassa sopravvivenza, scarsa integrazione e conservazione modesta nel tessuto danneggiato5. Inoltre, queste strategie basate sulle cellule non riescono a ripristinare a distanza axonal tratti, soprattutto in un modo controllato. Di conseguenza, biomateriali in combinazione con altri approcci stanno esplorande come veicoli di consegna per vari neurale e fattori di crescita e cellule progenitrici26. Approcci basati su biomateriali presentano un elevato grado di controllo di progettazione per la produzione di costrutti che imitano le specifiche fisiche, haptotaxic, e chemotaxic spunti presentano nel microambiente tridimensionale (3D) di destinazione host tessuto27, 28,29,30,31,32,33,34. Riproduzione di questi segnali ambientali è preminente per cellule trapiantate a presentare nativo-come la morfologia, la proliferazione, migrazione e segnalazione, tra le altre caratteristiche neurobiologiche29. Nonostante queste proprietà vantaggiose, avanzamento oltre tradizionale delle cellule seminate biomateriale ponteggi è necessaria per promuovere la rigenerazione assonale interurbano diretta e sostituire i neuroni persi contemporaneamente.
Un promettente approccio alternativo si basa su tessuto neurale ingegnerizzato “ponteggi vivente”, che sono distinti da altri approcci basati su cellule a causa della presenza di cellule neurali viventi con un cytoarchitecture preformato che emula neuroanatomia nativo e/o meccanismi per facilitare la sostituzione mirata, la ricostruzione e la rinascita di circuiti neurali4,35. Considerazioni per la progettazione di ponteggi viventi includono i fenotipi e le fonti di cellule neurali, come pure le proprietà fisiche e meccaniche e i segnali biochimici dettata dalla composizione di qualsiasi accompagnamento di biomateriali35. Dopo il montaggio in vitro, queste impalcature vivente possono essere impiantato in vivo di molecole di adesione cellulare presente e chemiotattica e neurotrophic segnali per regolare attivamente la migrazione delle cellule neurali e crescita assonale a seconda dello stato e progressione dei processi rigenerativi35. Le cellule gliali possono servire come base per i derivati dal cytoarchitecture di vita ponteggi poiché queste cellule mediano vari meccanismi in vivo. Durante lo sviluppo cerebrale, nuovi neuroni si basano su processi basali esteso da glia radiale dalla zona ventricolare verso la piastra corticale in via di sviluppo come vivente impalcature per migrazione diretto36,37. Inoltre, estendendo la crescita coni sono mostrato a orientarsi rilevando segnali attraenti e repellenti suscitati dalle cellule gliali guidepost e cosiddetto “pionieristico” assoni sono consigliati per raggiungere gli obiettivi corretti estendendo lungo pre-modellato glial ponteggi di35,38,39. Così, le cellule gliali sono necessari per la Guida di avanguardia di assoni, che più tardi servono come assone-based “ponteggi vivente” per dirigere la proiezione degli assoni “seguace”. Inoltre, meccanismi di crescita mediata da cellule gliali sono stati indicati a persistere dopo la nascita, come neuroblasti seguono migratori rostrale (RMS) per spostarsi dalla zona subventricolare (SVZ), una delle poche regioni di neurogenesi nel cervello adulto, per il bulbo olfattivo (OB)40. Questi neuroblasti nel RMS migrano all’interno del tubo glial (Figura 1A-1), che è costituito da processi astrocitari allineati longitudinalmente, tramite le adesioni cellula-cellula diretto e localizzato fattori solubili37, 41. Infine, mentre danno dello SNC in cause di mammiferi perturbato disposizione astrocytic processo formando una cicatrice gliale che fisicamente impedisce la rigenerazione assonale17, molti sistemi di mammiferi mancano la formazione di una cicatrice gliale dannosa. Piuttosto, le cellule gliali di specie di mammiferi mantengono più organizzato, allineato modelli che vengono utilizzati come guide attraverso la regione feriti17,42,43. Per esempio, nei modelli SCI non derivanti da mammiferi, assoni sono mostrati a crescere in stretta associazione con glial ponti che attraversano la lesione, suggerendo un ruolo importante per ponteggi glial organizzati come substrati facilitando la rigenerazione assonale e recupero funzionale ( Figura 1A -2) 42 , 44 , 45. ricapitolazione delle caratteristiche neuroanatomiche e i meccanismi dello sviluppo/rigenerativo descritti sopra può produrre una nuova classe di derivati dal vivere glial basato su impalcature che contemporaneamente possono guidare immatura migrazione neuronale ed assonale path-finding attraverso ambienti altrimenti non permissivi, quindi potenzialmente mitigare gli effetti di un neurone e la degenerazione del tratto dell’assone associato con lesione dello SNC e malattia.
Il nostro gruppo di ricerca ha già progettato più tipi di ponteggi vivente per la ricostruzione e rigenerazione di axonal tratti nel sistema nervoso centrale e sistema nervoso periferico (PNS) tramite micro-tessuto ingegnerizzato reti neurali (micro-teen) e tessuto innesti ingegnerizzati del nervo (TENGs), rispettivamente27,46,47,48. Entrambe le strategie dipendono intrinsecamente biomimetica. Micro-teen sono strutture anatomicamente ispirato alla strutturalmente e funzionalmente sostituire axonal tratti collegamento distinte popolazioni neuronali del cervello. TENGs sfruttare il meccanismo inerente allo sviluppo di assone-facilitato la rigenerazione assonale, esemplificato dalla crescita assonale “seguace” lungo gli assoni “pioniere”, per raggiungere la destinazione host la rigenerazione assonale35,46,48. Abbiamo recentemente sfruttato la versatilità dello scaffold vivente tecnica mediante uno schema simile di encasement come micro-teen e in cerca di ispirazione dai meccanismi basati su cellule gliali presenti nel corso dello sviluppo. Qui, abbiamo sviluppato costrutti costituito da fasci astrocytic allineati che abbracciano il lume collageno di un idrogel micro-colonna49. Queste impalcature astrocytic viventi sono state sviluppate da primo riempimento un gruppo di ago di agopuntura-tubo capillare con agarosio liquido per creare un idrogel cavo cilindrico con un diametro esterno (OD) e il diametro interno (ID) corrispondenti ai diametri della tubo ed ago, rispettivamente. A seguito dell’agarosi gelificazione ed estrazione di idrogel micro-colonna dal tubo capillare, interno cavo è rivestito con tipo I collagene per fornire un ambiente permissivo per adesione di astrociti e allineato bundle formazione (Figura 1B -1). In seguito, il lumen è seminato con i astrocytes corticali cerebrali isolati da cuccioli del ratto postnatale (Figura 1B-2). Contrariamente alle tecniche bidimensionali (2D) allineamento che si basano sull’applicazione di campi elettrici, bioerodibili scanalature e matrice extracellulare della proteina (ECM) patterning, allineamento di astrociti nell’impalcatura di vita tecnica si basa sulla auto-assemblaggio secondo variabili controllabili come substrato curvatura (ID di colonna), la densità delle cellule e collagene concentrazione50,51,52. Gli astrociti contraggono e rimodellano il collagene e di acquisiscono una morfologia bipolare, allineati longitudinalmente analoga per i ponteggi naturali osservati in vivo (Figura 1B-3). Infatti, stiamo attivamente perseguendo l’uso di questi cavo-come le strutture come substrati fisici per un orientamento mirato di migrazione neuroni immaturi, nonché facilitare la rigenerazione assonale attraverso l’ambiente sfavorevole del danneggiato snc, in particolare la cicatrice gliale mammiferi (Figura 1). Questo articolo verrà presentare il metodo di fabbricazione dettagliato per le micro-colonne astrocytic, contrasto e immunofluorescenza immagini della citoarchitettura previsto e per una discussione esaustiva sulle limitazioni attuali e future direzioni di fase i tecnica.
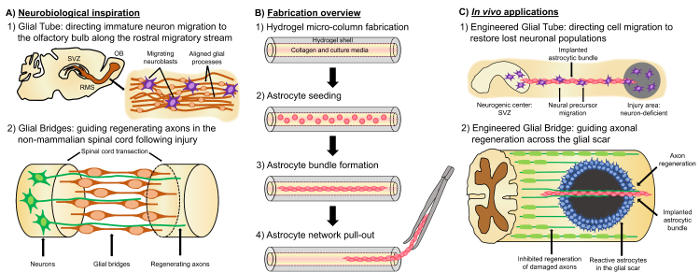
Figura 1: ispirazione, protocollo di fabbricazione e applicazioni proposte per le reti di Astrocytic allineate. Ispirazione neurobiologica (A): (1) neuroblasti provenienti dalla zona subventricular neurogena (SVZ) utilizzano il tubo glial allineato longitudinalmente in rostrale migratori (RMS) per la migrazione diretto verso il bulbo olfattivo (OB); (2) Non-mammiferi come anfibi e pesci possono sostenere la rigenerazione dopo tessuto neurale in parte i danni dovuti alla formazione di un ponte glial che collega le estremità di una lesione (ad es. transected del midollo spinale) e serve come impalcatura per la Guida di assoni di rigenerazione. (B) Panoramica di fabbricazione: (1) costruzione di un’idrogel micron di dimensioni medie, cavo micro-colonna con il lume rivestito con ECM, (2) semina di astrociti corticali primari isolati da cuccioli del ratto postnatale, (3) auto-assemblaggio di longitudinalmente orientati verso il fasci di cultura ed estrazione (4) del fascio dal encasement del biomateriale per gli studi futuri dell’impianto. (C) applicazioni In vivo : (1) Queste impalcature vivente possono servire come tubi gliali ingegnerizzati per migrazione neuronale diretto da neurogene centri per ripopolare le regioni del neurone-carenti; (2) ricapitolazione del meccanismo dello sviluppo di avanguardia di orientamento dell’assone e il meccanismo rigenerativo di glial ponti in non-mammiferi può dotare queste impalcature astrocitari con la capacità di dirigere la rigenerazione assonale attraverso il non permissivi ambiente della cicatrice gliale dei mammiferi. Clicca qui per visualizzare una versione più grande di questa figura.
