Центральной нервной системы (ЦНС) имеет ограниченные возможности противодействовать потери и/или дисфункции нейронов и аксональной пути, которые сопровождают условий, таких как черепно-мозговой травмы (ЧМТ), инсульт, травме (ТСТ) и нейродегенеративных заболеваний спинного1 ,–2,–3,–4,–5. Нейрогенез в ЦНС ограничивается ограниченное количество областей в мозге, препятствует восстановлению утраченных нейронов6,7. Кроме того восстановление утраченных аксональное путей в ЦНС недостаточно из-за отсутствия направленного указаний, присутствие ингибиторов нарост и реактивной astrogliosis после повреждения нервной ткани2,8, 9,10. Астроциты, как правило, имеют различные функции в деле оказания помощи нейронов с ионного гомеостаза, Распродажа нейромедиатора, формирования синапсов и нервно-сосудистых муфты11. Тем не менее после даже умеренные повреждения нервной ткани, астроциты могут пройти молекулярные, структурные и функциональные изменения, как они переход к гипертрофических состояние11. В ответ на серьезные нейротравма эти изменения приводят к формированию шрам с полутень, содержащие перенос реактивной астроциты и поражения ядро, которое включает в себя лейкоциты, утечка из расколовшегося гематоэнцефалический барьер (ГЭБ), микроглии, Олигодендроциты и фибробласты11,12,13. Эти реактивные астроциты достичь морфология нитевидные, неорганизованное процессов и демонстрируют увеличение выражение промежуточного накаливания белков и протеогликаны хондроитин сульфат (CSPGs), которые препятствуют нейронная регенерация12. Несмотря на то, что глиальный рубец первоначально помогает восстановить целостность BBB и избежать передачи воспалительной реакции на окружающие здоровые ткани, он служит физических и биохимических барьер против регенерации аксона12,14 ,,1516. Например аксоны которых сталкиваются глиальный рубец отображения луковичных дистрофические роста шишки и сдерживали рост12. Кроме того дезорганизация Астроцитарная процессов после травмы препятствует расширение регенерировать аксоны17. Итоги этих характеристик тормозной проявляется в часто постоянного физические и неврологические расстройства, которые пациенты страдают после тяжелой нейротравма, включая ЧМТ и SCI.
Независимо от того, внешние задачи, стоящие перед функциональным регенерации в ЦНС аксоны было показано, обладают внутреннюю способность к регенерации. Например динамический характер дистрофические роста конусов при контакте с глиальный рубец свидетельствует о том, что эти окончаний сохраняют свою способность расширять12. Следовательно считается, что основным препятствием для аксональное re рост является среды ингибирующих после травмы ЦНС и что предоставление более питательную среду через сокращение глиальных рубцов и/или предоставление восстановительной мосты через шрам будет выгодно. Действительно предыдущие исследования показали, что нейронов ЦНС способны расширения аксоны через поражения с помощью периферических нервов графтов как мосты, которые создают более благоприятные условия для axon регенерации12,18, 19. Ряд других стратегий предпринимались эксплуатировать этот рудиментарный регенеративной способностью. К примеру манипуляции клеток роста сигнальных путей в различных моделях травмы привело к аксональное регенерации и глиальных Шрам сокращения10,,2021. Кроме того исследования показали, что лечение с chondroitinase ABC, который расщепляет большинство цепочки сахара в CSPGs, уменьшает эффект CSPGs, выделяемый реактивной астроциты22. Несмотря на обнадеживающие результаты, эти подходы не дают направлен руководством конусы роста, которые потенциально может приводить к аномальным регенерации12, а также не учитывают потери нейронов. Подходы на основе клеток были использованы в попытках преодолеть последствия глиальный рубец и пополнить потерянные клеток, особенно нейронов. Некоторые группы Дедифференцированная реактивной астроциты в нейроны, в то время как другие пересажены нейронных прогениторных клеток в поражениях ЦНС населить области травмы и поощрять аксона регенерации23,24, 25. Однако, трансплантация стволовых клеток только ограничивается низкой выживаемости, плохое интеграции и скромный удержания в поврежденной ткани5. Кроме того эти стратегии на основе клеток удастся восстановить междугородной аксональное участки, особенно в контролируемым образом. Таким образом биоматериалов в сочетании с другими подходами в настоящее время изучаются как средства доставки для различных нейронных и клеток-предшественников и роста факторы26. Подходы на основе биоматериала имеют высокую степень управления дизайн для производства конструкций, которые имитируют конкретные физические, haptotaxic, и chemotaxic сигналы в трехмерной (3D) микроокружения целевого узла ткани27, 28,29,30,,3132,,3334. Воспроизведение этих экологических сигналов имеет первостепенное значение для пересаженные клетки представить родной как морфология, распространения, миграции и сигнализации, среди других нейробиологических характеристики29. Несмотря на эти выгодные свойства улучшению за пределами традиционных клеток посеян биоматериала подмостей требуется одновременно направлены междугородной аксональное регенерации и заменить потерянные нейронов.
Обещая альтернативный подход основан на нервной ткани инженерии «живые леса», которые отличаются от других подходов, основанных на ячейки, из-за присутствия живых нервных клеток с предварительно cytoarchitecture, что эмулирует родной нейроанатомия и/или развития механизмов для облегчения целевых замена, восстановление и регенерация нейронной схемы4,35. Соображения по проектированию жизни леса включают фенотипы и источники нервные клетки, а также механические/физические свойства и биохимические сигналы продиктовано состав любых сопровождающих биоматериалов35. После изготовления в пробирке, эти живые леса может быть имплантирован в естественных условиях настоящего молекул клеточной адгезии и эозинофилов и нейротрофическими сигналы активно регулировать миграцию нервных клеток и аксон нарост в зависимости от государства и прогрессирование регенераторные процессы35. Глиальные клетки может служить основой для инженерных cytoarchitecture жизни леса, так как эти клетки посредником различные механизмы развития в естественных условиях. Во время развития мозга новые нейроны полагаются на базальную процессы, продлен на радиальной глии из желудочков зоны к развивающихся корковых пластины как живые леса для миграции направлены36,37. Кроме того расширение роста шишки показано ориентироваться по зондирования привлекательным и репелленты сигналы с ориентиром глиальных клеток и так называемые «первопроходца» аксоны предлагается достичь правильной цели, расширяя вдоль заранее узорной глиальных помостами35,,3839. Таким образом, глиальные клетки необходимы для руководства первопроходца аксонов, которые впоследствии служат как на основе аксон «живые леса» в прямой проекции «последователь» аксоны. Кроме того, механизмы роста глии опосредованной было показано постнатально, сохраняются как нейробласты следовать ростральной миграционных потока (RMS) для перехода из субвентрикулярной зоны (SVZ), один из немногих оставшихся областей нейрогенез взрослого мозга, обонятельные луковицы (OB)40. Эти нейробласты в RMS мигрируют в глиальных трубки (рис. 1а-1), который состоит из продольно в соответствие Астроцитарная процессов, через прямой ячеек спаек и локализованные растворимых факторов37, 41. Наконец, хотя повреждения ЦНС в млекопитающих причин нарушается механизм Астроцитарная процесса формирования глиальных шрам, который физически препятствует аксональное регенерации17, многие системы не млекопитающих не хватает формирования глиальных наносит шрам. Скорее глиальные клетки не млекопитающих видов поддерживать более организованной, соответствие шаблонов, которые используются в качестве руководства через пострадавшего региона17,42,43. Например в моделях SCI не млекопитающих, аксоны показываются расти в тесном сотрудничестве с глиальных мосты, пересекая поражения, предлагая важную роль для организованных глиальных леса как субстратов, содействия аксональное регенерации и восстановления функций ( Рисунок 1A -2) 42 , 44 , 45. перепросмотре нейроанатомический функций и развития/восстановительных механизмов, описанных выше может принести новый класс инженерии глиальных основе жизни леса, которые могут одновременно управлять незрелых нейронной миграции и аксональной Поиск пути через иначе либеральной среды, тем самым потенциально смягчения последствий нейронов и аксон тракта дегенерации, связанные с ЦНС травмы и болезни.
Наша исследовательская группа ранее разработала несколько типов живых подмости для восстановления и регенерации аксональное участки в ЦНС и периферической нервной системы (ПНС) через микро ткани спроектирован нейронных сетей (микро TENNs) и ткани графтов инженерии нерва (TENGs), соответственно27,46,47,48. Обе стратегии по существу основаны на biomimicry. Микро-TENNs являются анатомически вдохновил сооружения, предназначенные для структурно и функционально заменить аксональное участки, подключение различных нейрональных популяций головного мозга. TENGs использовать механизму развития аксон способствовали аксональное регенерации, подтверждается «последователь» аксон роста вдоль аксоны «Пионер», для достижения целевой хост аксональное регенерации35,46,48. Мы недавно капитализируются на универсальность живых эшафот техника с использованием аналогичной схемы оболочки как микро TENNs и ищет вдохновение от механизмов на основе глии представить на протяжении развития. Здесь мы разработали конструкции, состоящей из унифицированных Астроцитарная связки, охватывающих коллагеновых люмен гидрогеля микро колонки49. Эти леса Астроцитарная жизни, разработанные Первое заполнение сборку капиллярной трубки иглоукалывание иглы с жидким агарозы для создания полые цилиндрические гидрогеля с внешнего диаметра (OD) и внутренний диаметр (ID) соответствующих диаметров трубки и иглы, соответственно. После геля агарозы и извлечение микро столбце гидрогеля с капиллярной трубки, полые интерьер является покрытием с типом коллаген предоставлять среде разрешительных для экзоцитоз адгезии и соответствие комплекта формирования (рис. 1B -1). После этого просвет заполняется с мозговой коры астроциты, изолированы от послеродового крыса щенков (Рисунок 1B-2). Вопреки двухмерный (2D) выравнивания методы, которые полагаются на применение электрических полей, micropatterned канавки и внеклеточного матрикса (ECM) белка кучность, экзоцитоз выравнивания в жизни леса способ полагается на самостоятельной сборки по данным контролируемых переменных, таких как субстрат кривизны (столбец ID), плотность ячеек и коллаген концентрации50,51,52. Астроциты контракт и перестроить коллагена и приобрести биполярный, продольно соответствие морфологии, аналог природных лесов, наблюдается в vivo (рис. 1B-3). Действительно мы активно осуществляем использование этих кабель подобных структур как физические субстратов для целенаправленного руководства миграции незрелых нейронах, а также содействия аксональное регенерации в неблагоприятной окружающей среде повреждения ЦНС, особенно млекопитающих глиальный рубец (рис. 1 c). Эта статья представит подробные изготовления для Астроцитарная микро колонки, фаза контраст и иммунофлюоресценции изображения ожидаемого cytoarchitecture и всеобъемлющее обсуждение текущих ограничений и будущие направления деятельности техника.
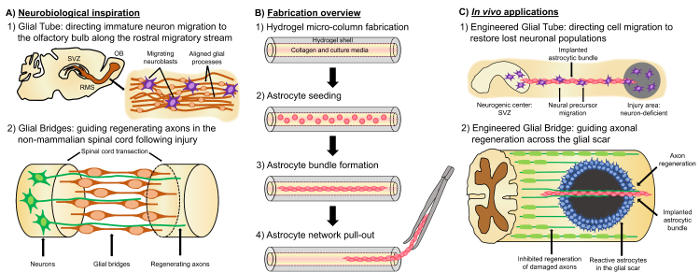
Рисунок 1: вдохновение, изготовление протокол и предлагаемых приложений для выровненного Астроцитарная сетей. (A) нейробиологических вдохновение: (1) нейробласты возникая от нейрогенный субвентрикулярной зоны (SVZ) использовать продольно унифицированных глиальных трубки в ростральной миграционных потока (RMS) для направленного миграции к обонятельной луковицы (OB); (2) Non млекопитающих, таких как амфибий и рыб могут поддерживать регенерации после нервной ткани ущерб отчасти за счет формирования глиальных мост, который соединяет заканчивается поражением (например пересекал спинного мозга) и служит леску для руководства регенерирующее аксоны. (B) Обзор изготовления: (1) строительство микро столбца микронного размера, полые гидрогеля с люмен, покрытые ECM, (2) заполнение первичной коры астроциты, изолированы от послеродового крыса детенышей, (3) самосборки продольно ориентированных связки в культуре и (4) добыча комплект из оболочки биоматериала для исследования будущей имплантации. (C) в естественных условиях применения: (1) эти живые леса могут служить инженерии глиальных трубы для направленного нейрон миграции из нейрогенный центров, чтобы населить нейрон недостаточным регионов; (2) резюме развития механизма новаторской руководство аксона и регенеративной механизм глиальных мостов в не млекопитающих может наделить эти Астроцитарная леса с способностью прямой регенерации аксона через не разрешительной окружающей среды млекопитающих глиальный рубец. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
