Sistema nervioso central (SNC) tiene una capacidad limitada para contrarrestar la pérdida o disfunción de las neuronas y vías axonales que acompañan las condiciones tales como lesión cerebral traumática (TBI), movimiento, médula espinal lesión (SCI) y neurodegenerative enfermedad1 ,2,3,4,5. Neurogénesis en el SNC se limita a un número limitado de áreas en el cerebro, dificultando la restauración de las neuronas perdidas6,7. Además, la regeneración de caminos perdidas axonales en el SNC es insuficiente debido a la falta de orientación dirigida, la presencia de inhibidores de la consecuencia y astrogliosis reactiva después de daño al tejido neural2,8, 9,10. Los astrocitos tienen diversas funciones para ayudar a las neuronas con la homeostasis de iones, separación de neurotransmisores, formación de sinapsis y neurovasculares acoplamiento11. Sin embargo, siguiendo incluso leves daños de tejido neural, astrocytes pueden sufrir cambios moleculares, estructurales y funcionales en su transición a un estado hipertrófico11. En respuesta al neurotrauma grave, estos cambios resultan en la formación de una cicatriz con una penumbra que contienen astrocitos reactivos migrando y un núcleo de la lesión que incluye leucocitos se filtró desde el rota barrera blood – brain (BBB), microglia, oligodendrocitos y fibroblastos11,12,13. Estos astrocitos reactivos logran una morfología de procesos filamentosos, desorganizados y presentan aumento de la expresión de proteínas de filamentos intermediarios y proteoglicanos de sulfato del chondroitin (CSPGs), que impiden la regeneración de los nervios12. A pesar de la cicatriz glial inicialmente ayuda a restaurar la integridad de la BBB y evitar la transmisión de la respuesta inflamatoria al tejido sano circundante, sirve como una barrera física y bioquímica contra axon regeneración12,14 ,15,16. Por ejemplo, axones que encuentran la cicatriz glial mostrar los conos de crecimiento distrófico con bulbo y retraso de crecimiento12. Además, la desorganización de los procesos astrocytic después de la lesión impide la extensión de regenerar axones17. El resultado de estas características inhibitorias se manifiesta en las debilitaciones físicas y neurológicas a menudo permanente que los pacientes sufren después de neurotrauma grave, incluyendo TBI y la médula espinal.
Independientemente de los retos extrínsecos de regeneración funcional en el SNC, axones han demostrado poseer una capacidad intrínseca de regeneración. Por ejemplo, la naturaleza dinámica de los conos de crecimiento distrófico en contacto con la cicatriz glial sugiere que estas terminaciones conservan su capacidad para ampliar a12. En consecuencia, se cree que un obstáculo principal para el crecimiento axonal es el entorno inhibidor del SNC después de la lesión y proporciona un ambiente más permisivo a través de reducción de la cicatrización glial o proporcionar puentes regenerativas a través de la cicatriz sería ventajoso. De hecho, estudios anteriores han demostrado que las neuronas de la CNS fueron capaces de extender axones a través de una lesión mediante injertos de nervios periféricos como puentes, que presentan un entorno más favorable para la regeneración de axón12,18, 19. Varias otras estrategias han sido llevadas a cabo para explotar esta capacidad regenerativa vestigial. Por ejemplo, manipulación de vías de señalización de crecimiento celular en varios modelos de lesión ha dado lugar a la regeneración axonal y disminución de cicatriz glial10,20,21. Además, los estudios han demostrado que el tratamiento con Condroitinasa ABC, que hiende la mayoría de las cadenas de azúcar en CSPGs, disminuye el efecto inhibitorio de CSPGs segregada por los astrocitos reactivos22. A pesar de alentador resultados, estos métodos no proporcionan dirigida orientación de conos de crecimiento, que potencialmente puede resultar en la regeneración aberrante12y también no tienen en cuenta para la pérdida de neuronas. Enfoques basados en la célula se han utilizado en tentativas de superar los efectos de la cicatriz glial y para reponer las células perdidas, particularmente de las neuronas. Algunos grupos han dedifferentiated astrocitos reactivos en las neuronas, mientras que otros han trasplantado células progenitoras neurales en lesiones CNS para repoblar la zona de lesión y promover axon regeneración23,24, 25. sin embargo, la célula de vástago trasplante solo está limitado por las tasas de supervivencia bajas, pobre integración y retención modesta en el tejido dañado5. Además, estas estrategias basadas en células incapaces de restaurar zonas interurbana axonal, especialmente en forma controlada. Por lo tanto, biomateriales en combinación con otros enfoques se están estudiando como vectores para varios nervios y factores de crecimiento y células progenitoras26. Enfoques basados en el biomaterial cuentan con un alto grado de control del diseño para producir construcciones que imitan a la haptotaxic física, específica, y chemotaxic señales presentan en el microambiente (3D) tridimensional del destino anfitrión tejido27, 28,29,30,31,32,33,34. Reproducción de estas señales ambientales es de suma importancia para las células trasplantadas presentar nativo-como morfología, proliferación, migración y señalización, entre otras características neurobiológicas29. A pesar de estas propiedades ventajosas, avance más allá de la tradicional células sembradas biomaterial andamios es necesario simultáneamente promover la regeneración axonal larga distancia dirigida y reemplazar las neuronas perdidas.
Un prometedor enfoque alternativo se basa en el tejido neural ingeniería “andamios de vida”, que son distintos de otros enfoques basados en la célula debido a la presencia de células neuronales vivas con una Citoarquitectura preformado que emula neuroanatomía nativa o desarrollo mecanismos que faciliten la sustitución específica, reconstrucción y regeneración de los circuitos neuronales4,35. Consideraciones para el diseño de matrices de soporte de vida incluyen los fenotipos y las fuentes de las células neuronales, así como las propiedades físico mecánicas y las señales bioquímicas dictaron por la composición de cualquier adjunto de biomateriales35. Después de la fabricación en vitro, estos andamios de vida pueden ser implantados en vivo quimiotácticos y moléculas de adhesión celular presentes y neurotróficos señales para regular activamente migración de células neuronales y consecuencia del axón dependiendo del estado y progresión de procesos regenerativos35. Las células gliales pueden servir como base para la ingeniería Citoarquitectura de andamios de la vida ya que estas células median varios mecanismos del desarrollo en vivo. Durante el desarrollo del cerebro, las neuronas nuevas dependen de procesos basals extendidos por glia radial desde la zona ventricular hacia la placa cortical en vías de desarrollo como andamios de vida migración dirigida36,37. Además, extender crecimiento conos son muestra a orientarse detectando señales atractivas y repulsivo provocadas por las células gliales hito y supuesto “pioneras” axones son sugeridas para alcanzar los objetivos correctos por que se extiende a lo largo de los estampados glial Andamios de38,35,39. Así, las células gliales son necesarias para la dirección del pionero axons, que luego sirven como base de axon “andamios de vida” para dirigir la proyección de axones “seguidor”. Por otra parte, mecanismos de crecimiento mediada por la glía han demostrado persistencia postnatal, como neuroblastos siguen la corriente migratoria rostral (RMS) para poder navegar desde la zona subventricular (SVZ), una de las pocas áreas restantes de la neurogénesis en el cerebro adulto, la bulbo olfatorio (OB)40. Estos neuroblastos en el RMS migran dentro del tubo de glial (figura 1A-1), que se compone de procesos astrocytic alineadas longitudinalmente, mediante adhesiones célula-célula directa y localizada de factores solubles37, 41. por último, mientras que el daño del CNS en causas de mamíferos interrumpió arreglo astrocytic proceso formando una cicatriz glial que físicamente impide la regeneración axonal17, muchos sistemas no mamíferos carecen de la formación de una cicatriz glial perjudicial. Más bien, las células gliales de las especies de animales no mamíferos mantienen más organizado, alineados los patrones que se utilizan como guías a través de la región lesionada17,42,43. Por ejemplo, en modelos de animales no mamíferos SCI, se muestran axones que crecen en estrecha asociación con puentes gliales cruzar la lesión, lo que sugiere un papel importante para andamios gliales organizados como sustratos facilitando la regeneración axonal y recuperación funcional ( Figura 1A -2) 42 , 44 , 45. recapitulación de las características neuroanatómicas y los mecanismos del desarrollo/regenerativa descritos anteriormente puede producir una nueva clase de andamios de ingeniería vida glial base que puede conducir al mismo tiempo la migración neuronal inmadura y axonal pathfinding a través de entornos no permisiva, de tal modo potencialmente mitigar los efectos de neuronal y degeneración de la zona de axón asociado a enfermedad y lesión del CNS.
Nuestro grupo de investigación previamente ha diseñado varios tipos de matrices de soporte de vida para la reconstrucción y regeneración del tracto axonal en el SNC y sistema nervioso periférico (SNP) a través de micro-tejido ingeniería de redes neuronales (micro-adolescentes) y tejido Ingeniería injertos del nervio (TENGs), respectivamente27,46,47,48. Ambas estrategias están basados inherentemente en biomimética. Micro-adolescentes son estructuras anatómicamente inspiración estructural y funcionalmente sustituye extensiones axonales conectar distintas poblaciones neuronales del cerebro. TENGs explotan el mecanismo del desarrollo de la regeneración axonal facilitado por axon, ejemplificado por el crecimiento del axon de “seguidor” a lo largo de axones de “pionero”, para lograr la regeneración axonal de host destino35,46,48. Nos recientemente aprovechó la versatilidad de la matriz de soporte de vida técnica mediante un esquema similar de protector como micro-adolescentes y buscando inspiración en los mecanismos basados en la glia presentes en todo el desarrollo. Aquí, hemos desarrollado construcciones consisten en paquetes astrocytic alineados que atraviesan el lumen colagenoso de una micro columna de hidrogel49. Estos andamios vida astrocíticos son desarrollados por el primer llenado de un conjunto de agujas de acupuntura de tubo capilar con agarosa líquida para crear un hidrogel cilíndrico hueco con un diámetro exterior (OD) y diámetro interno (ID) correspondiente a los diámetros de la tubo y aguja, respectivamente. Gelificación de agarosa y extracción de la hidrogel micro-columna de tubo capilar, el hueco interior es revestido con tipo y colágeno para suministrar un entorno permisivo para la adherencia de astrositos y alineado paquete formación (figura 1B -1). Luego, la luz es sembrada con astrocitos corticales cerebrales aislados de crías de rata postnatal (figura 1B-2). Contrariamente a las técnicas bidimensionales (2D) alineación que se basan en la aplicación de campos eléctricos, micropatterned surcos y matriz extracelular proteína (ECM) patrones, alineación de astrositos en el andamio de la vida técnica depende de uno mismo-Asamblea de acuerdo a las variables controlables como curvatura de sustrato (columna ID), la densidad celular y la concentración de colágeno50,51,52. Los astrocitos contratan y remodelan el colágeno y adquieran una morfología bipolar, alineados longitudinalmente análoga a las matrices de soporte naturales en vivo (figura 1B-3). De hecho, estamos persiguiendo activamente el uso de estas estructuras de cable como sustratos físicos específicos guía de migración de las neuronas inmaduras, así como facilitar la regeneración axonal a través del ambiente desfavorable del SNC dañado, particularmente la cicatriz glial mamífera (figura 1). Este artículo se presenta el método de fabricación detallado para las columnas de micro astrocytic, fase imágenes de contraste, inmunofluorescencia y de la Citoarquitectura esperado y una amplia discusión sobre las limitaciones actuales y direcciones futuras de la técnica.
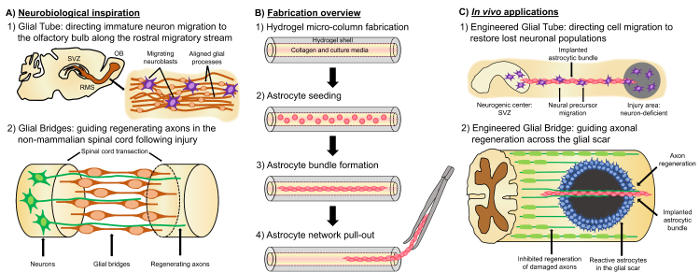
Figura 1: inspiración, protocolo de fabricación y aplicaciones propuestos para las redes Astrocytic alineadas. (A) inspiración neurobiológica: neuroblastos (1) origina la neurogénica zona subventricular (SVZ) utilizan el tubo glial longitudinalmente alineado en la corriente migratoria rostral (RMS) para la migración dirigida hacia el bulbo olfatorio (OB); (2) no de mamíferos como peces y anfibios pueden sostener la regeneración después de tejido neural dañar en parte debido a la formación de un puente glial que conecta los extremos de una lesión (por ejemplo médula espinal transected) y sirve como un andamio para la dirección de regeneración de axones. (B) Resumen de fabricación: (1) construcción de una micro columna hidrogel de tamaño micrométrico, hueco con recubierta de ECM, la luz de la (2) siembra de astrocitos corticales primarios aislados de crías de rata postnatal, (3) uno mismo-Asamblea de longitudinalmente orientados paquetes en la cultura y (4) extracción del paquete del encasement del biomaterial para estudios de implantación futura. (C) In vivo usos: (1) estos andamios de vida pueden servir como tubos gliales ingeniería para la migración de la neurona dirigida de centros neurogénicas a repoblar las regiones deficientes en neurona; (2) Resumen del mecanismo del desarrollo de pioneras orientación axón y el mecanismo regenerativo de puentes gliales en los mamíferos no puede dotar a estos andamios astrocytic con capacidad de dirigir la regeneración del axón a través de la no-permisivo medio ambiente de la cicatriz glial mamífera. Haga clic aquí para ver una versión más grande de esta figura.
