Микроорганизмы, связанные с растениями, играют важную роль в биогеохимическом циклировании, биоремедиации, смягчении последствий изменения климата, роста растений и здоровья и устойчивости растений к биотическим и абиотическим стрессам. Микроорганизмы взаимодействуют с растениями как непосредственно через контакт стенки стенки растения, так и косвенно через химическую секрецию и сигнализацию 1 , 2 , 3 . В качестве сидячих организмов растения разработали прямые и косвенные механизмы противодействия инфекции патогенами. Прямые защитные средства включают структурную защиту и экспрессию защитных белков, тогда как косвенная защита включает в себя вторичное производство метаболитов растений и привлечение организмов, антагонистических к вторгающимся патогенам 4 , 5 . Вырожденные корневым экссудатом, выделениями, слизи, мусигелем и лизатами изменяют физико-химические свойства ризосферы для привлечения или отталкиванияМикробов к их хозяевам 6 . Химический состав секреции корней является видоспецифичным, тем самым выступая в качестве селективного фильтра, который позволяет некоторым микроорганизмам, способным распознавать такие соединения, процветать в ризосфере 6 . Таким образом, совместимые микробные виды могут стимулироваться для активации и усиления их ассоциаций либо в пользу, либо в ущерб хозяину растения 1 .
Понимание взаимодействия растений и микробов в ризосфере является ключевым фактором повышения производительности растений и функционирования экосистем, поскольку большинство микробных и химических воздействий происходит в корневой структуре и почвенно-воздушном интерфейсе 2 , 6 , 7 , 8 . Тем не менее, изучение взаимодействия подземных растений и микробов и ответных реакций было проблемой из-за его интригующей Сложный и динамичный характер и отсутствие подходящих экспериментальных моделей с естественной структурой корней и морфологией растений в условиях жестко контролируемого роста. В качестве одного из наиболее интенсивно изучали фитопатогенов, Agrobacterium заражает широкий ассортимент растений с сельскохозяйственной и садоводческой значение, в том числе вишни, яблони, груши, виноград и розы 9. Agrobacterium – важный модельный организм для понимания взаимодействия растений с патогенами и является мощным инструментом в трансформации растений и инженерии растений 10 , 11 , 12 , 13 , 14 .
Молекулярное растение-взаимодействие Agrobacterium хорошо изучено в течение нескольких десятилетий, а современное понимание патогенности Agrobacterium обширно 9 ,F "> 11 , 15 , 16. Патогенность Agrobacterium в значительной степени объясняется его развитыми возможностями восприятия сигналов, полученных из растений, что приводит к тонкой модуляции его программы вирулентности и связи между клетками, так называемому восприятию кворума 17 . Программа вирулентности Agrobacterium регулируется несколькими сигналами, доступными в ризосфере, и включает в себя два набора 2-компонентных систем, систему ChvG / I и систему VirA / G. Кислотные условия в ризосфере активируют транскрипцию chvG / I , virA / G , И несколько других генов, участвующих в патогенезе Agrobacterium , включая virE0 , virE1 , virH1 , virH2 и гены системы секреции типа VI (T6SS) 18. Фенольные соединения растительного происхождения, включая ацетозиригон (4'-гидрокси-3 ', 5 '-диметоксиацетофенон), активировать VIrA / G 2-компонентная система через механизмы сигнализации фосфорилирования 19 . Затем VirA / G активирует весь регулятор vir , что приводит к переносу и интеграции бактериального фрагмента ДНК размером 20 кБ, называемого передаточной ДНК (Т-ДНК) из его индуцирующей опухоль (Ti) плазмиды в ядро растения 16 . Т-ДНК несет гены, ответственные за синтез растительных гормонов индол-3-уксусной кислоты (IAA) ( iaaM и iaaH ) и цитокинина ( ipt ), и один раз экспрессируется в растительных клетках, образуется большое количество этих фитогормонов. Это приводит к аномальной пролиферации тканей и развитию опухоли растений, известной как болезнь коронального желчного пузыря, что является хронической и нерегулярной проблемой для растений 9 , 11 , 20 . IAA также действует совместно с салициловой кислотой и гамма-аминомасляной кислотой, чтобы подавить вирулентность Agrobacterium или уменьшить Agrobacteriu М кворум (QS) 17 , 21 , 22 . Чтобы противостоять этой репрессии, Т-ДНК также несет гены для биосинтеза опиона, который активирует чувствительность кворума Agrobacterium, чтобы способствовать патогенности Agrobacterium, а также служит источником питательных веществ для патогена 22 , 23 .
Несмотря на полное глубокое понимание взаимодействия Agrobacterium- plant и результирующей передачи Т-ДНК в хозяин растения, сложные сигнальные события на начальной стадии взаимодействия менее понятны. Это частично связано с ограничениями обычных подходов к исследованию сигнализации Agrobacterium- plant. Культуры суспензии клеток растений и искусственное рандование, специфичное для конкретного участка, обычно используются для изучения молекулярных взаимодействий с микроорганизмами 24 ,Ef "> 26 , 27. Однако клеточные суспензии не имеют типичной морфологии растений, в частности, суспензионные клетки растений не имеют корневых структур и корневых экссудатов, которые очень важны для активации микробного хемотаксиса и вирулентности 28 , 29. Содержание морфологии растений И корневая структура была устранена искусственно ранившимися растениями, что облегчает сайт-специфическую инфекцию, что приводит к обнаружению индуцированных генов, связанных с защитой растений, в непосредственно инфицированной растительной ткани 30 , 31. Однако искусственное ранение значительно отличается от патогенной инфекции в природе , В частности, поскольку ранение приводит к накоплению жасмоновой кислоты (JA), которое системно препятствует передаче сигналов естественного растения и защите 26. Кроме того, синтетические химикаты обычно используются для искусственного стимулирования реакции хозяина растенийИли вирулентности патогена. Хотя возможно добавление таких химических соединений, отражающих концентрацию в плазме, такое добавление не учитывает диффузию корневых экссудатов постепенно в окружающую ризосферу, которая генерирует хемотаксический градиент, определяемый микробами 28 , 32 . Учитывая ограничения обычных подходов к изучению взаимодействий растений и микробов, точность и глубина полученных данных могут быть затруднены и ограничены, а знания, полученные из обычных подходов, могут не переводить непосредственно в planta . Многие аспекты сигнальной передачи растений – Agrobacterium еще не полностью поняты, особенно на ранней стадии взаимодействия, когда симптомы болезни еще не развиты.
Чтобы изменить ограничения обычных подходов, эта работа представляет собой недорогую, жестко контролируемую и гибкую гидропонику cКоторая позволяет исследователям глубже проникнуть в сложные сигнальные и ответные пути на начальном этапе взаимодействия молекулярных растений с микробами. Гидропоника широко используется для изучения питательных веществ растений, корневых экссудатов, условий роста и воздействия металлической токсичности на растения 33 , 34 . Существует несколько преимуществ гидропонных моделей, в том числе небольшие пространственные требования, доступность различных растительных тканей, жесткий контроль над питательными / экологическими условиями и борьба с вредителями / болезнями. Гидропонические системы также менее ограничивают рост растений по сравнению с методами агара / фитогара, которые обычно ограничивают рост через 2-3 недели. Важно отметить, что содержание цельнозерновых структур способствует естественной корневой секреции, необходимой для микробного хемотаксиса и индукции вирулентности 8 , 29 . Система descriПостель здесь проще и менее трудоемко, чем альтернативы 33 , 34 . Он использует меньше деталей и не требует каких-либо инструментов, кроме стандартных ножниц. Он использует металлическую сетку (в отличие от нейлона 33 ) в качестве сильной поддержки роста растений и простой метод аэрации в стерильных условиях путем встряхивания для поддержки роста микроорганизмов. Кроме того, система может использовать металлическую сетку различных размеров для поддержки роста растений, в которой размещаются разнообразные виды растений, не ограничивая ширину их корней.
В представленной здесь системе гидропонной кокультивации растения выращивают в стерильной гидропонной системе, где корни растений выделяют органические соединения, поддерживающие рост инокулированных бактерий. В этой системе кокультивации никакие искусственные химические вещества, такие как растительные гормоны, защитный элиситор или вызывающие вирулентность химические вещества, не дополняются, что отражает естественную клетку-сигнальный гомеостаз во время взаимодействия растений с микробами. С помощью этой гидропонной системы кокультивации можно было одновременно определить экспрессию гена в корневой ткани Arabidopsis thaliana Col-0 при инфицировании Agrobacterium , а также активацию генов Agrobacterium при кокультивации с Arabidopsis . Кроме того, было продемонстрировано, что эта система подходит для изучения присоединения Agrobacterium к корням растений, а также к профилю секреторного корня растения, при кокуляции (инфекции) Agrobacterium ( рис. 1 ).
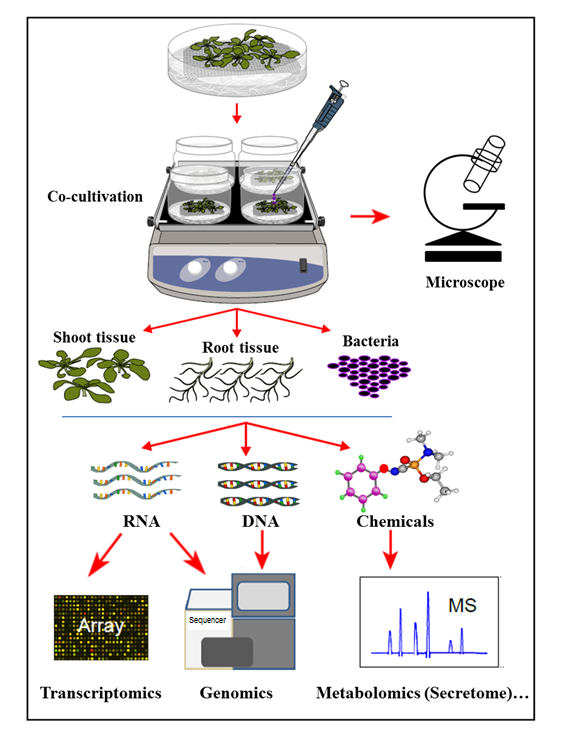
Рисунок 1: Обзор системы гидропонной кокультивации с анализом проб. Растения выращивают поверх сетки (побеги над сеткой), причем корни погружаются в гидропонную среду, которая затем инокулируется бактериями fИли сокультура. Затем растительные ткани и бактерии разделяют для одновременных экстракций и анализов. Эта цифра была изменена из ссылки 35 .
