The objective of this study was to assess the spatiotemporal expression pattern of nodule-specific P. vulgaris NIN. To do this, a 700 bp region upstream of the translation initiation codon of the NIN gene was selected and a set of oligos was designed as depicted in Figure 1A. Using a high-fidelity polymerase, the NIN promoter fragment was amplified, isolated (Figure 1B), and cloned into the destination binary vector pBGWFS7.0 (Figure 1C) through a Gateway approach to generate a transcriptional fusion to the chimeric reporter GUS-enhanced GFP (pBGWFS7.0/PvNIN::GUS-GFP). The pBGWFS7.0/PvNIN::GUS-GFP vector construct typically generates hundreds of independent clones. Colony screening by PCR with gene-specific oligos easily identifies the correct clones (Figure 2D). The PCR positive plasmid was Sanger sequenced with the promoter-specific oligos to verify the authenticity of the insert.
Maintaining high humidity throughout the experiment is necessary for the induction of callus and hairy roots. At the site of wounding, calli are usually observed at 5 to 7 days (Figure 2J) and ≥ 2 cm hairy roots are observed at 10 to 12 days after transformation. The majority of A. rhizogenes transformed plants successfully to produce hairy roots by 2 weeks (Figure 2K). However, the number and length of hairy roots can be variable. Therefore, select the most vigorously growing roots for analysis and further experiments. Excise the primary root by cutting the stem 2 cm below the hairy roots and transplant them into pots containing sterile vermiculite (Figure 2L) or in a hydroponics system.
To study the spatiotemporal expression of the nodule-specific NIN promoter, hairy roots were inoculated with R. tropici, and GUS activity was observed at periodic intervals post-inoculation depending on the experimental need. Inoculation with R. tropici induced a strong GUS expression in the transgenic roots, indicating that rhizobia induces the NIN expression in the common bean (Figure 3A). Further, sampling at 6-9 dpi showed GUS expression in Rhizobium infected root hair cells (Figure 3B). Phaseolus nodule primordium, and young and mature nodules also demonstrated NIN expression (Figure 3C). At all stages of nodulation, no such GUS expression was seen in the vector control roots (Figure 3D, 3E, 3F).
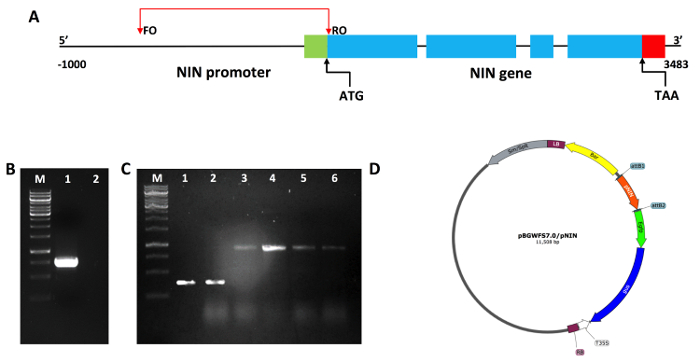
Figure 1: Outline of P. vulgaris NIN gene structure and cloning. (A) A schematic representation of the NIN gene structure showing 5'UTR (green), 4 exons, 3 introns, and 3'UTR (red) on chromosome number 9 of P. vulgaris. Upstream to the NIN translation initiation codon ATG, shows a promoter region on which oligos were designed. FO, forward oligo; RO, reverse oligo; ATG, start codon; TAA, stop codon. (B) PvNIN promoter region was amplified from freshly isolated P. vulgaris genomic DNA using the promoter-specific oligo set. M, 1 Kb ladder; 1, PCR reaction with gDNA showing amplicon size of 700 bp; 2, PCR reaction without gDNA (-ve control). (C) Screening of the pENTR/D-TOPO-PvNIN plasmids by PCR using the vector specific M13 oligos. Correct insertions have a band at 1,024 bp (lane 3-6) and vectors without inserts will have a 324 bp band (lane 1-2). (D) Promoter expression binary vector map used in the present study, pBGWFS7.0 vector showing PvNIN promoter in fusion with GUS and GFP. Please click here to view a larger version of this figure.
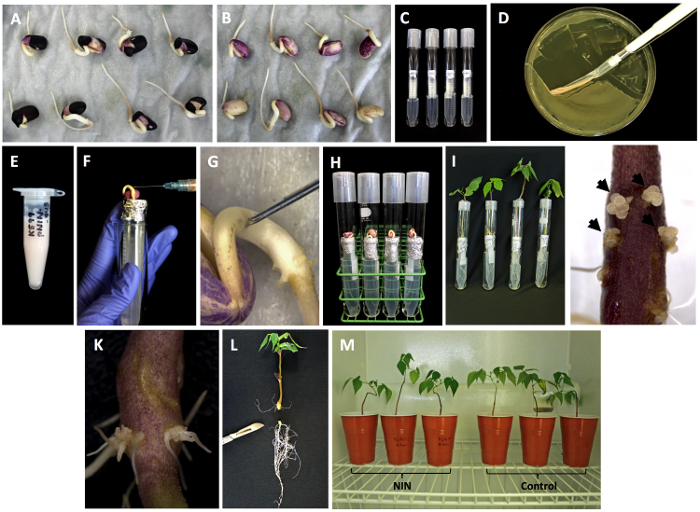
Figure 2: A. rhizogenes transformed hairy roots of the common bean. (A) The sterilized seeds of P. vulgaris cv. Negro Jamapa germinating on sterile filter paper. (B) Seeds, coat removed. (C) Tube setup for hairy root induction. (D) Scraping out A. rhizogenes culture (pNIN & control). (E) A. rhizogenes culture resuspended in sterile distilled water. (F, G) Injecting bacterial cells into hypocotyles. (H) Tube setup with Phaseolus seedlings injected with bacterial cells. (I, J) Calli formed at wounded site 5-7 dpi. (K) Hairy roots 15 dpi (L) excising the tap roots 2 cm below the site of hairy root induction. (M) Composite plants transplanted into sterile vermiculite inoculated with R. tropici. Please click here to view a larger version of this figure.
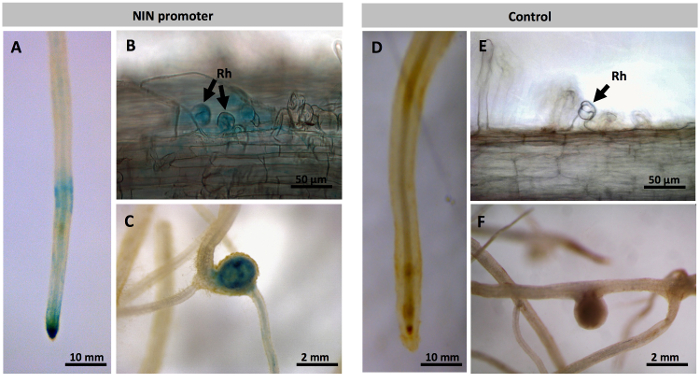
Figure 3: Promoter analysis of P. vulgaris NIN in transgenic P. vulgaris roots. Spatial and temporal pattern of NIN expression revealed by a promoterNIN::GUS-GFP construct in transgenic hairy roots incubated with GUS as a substrate. Optical microscope images of PvNIN:GUS : (A) Roots inoculated with R. tropici, (B) GUS expression in curled root hairs, and (C) young nodules. Images of control (empty vector) showing no such GUS expression in (D) roots inoculated with R. tropici, (E) curled root hairs, and (F) young nodules. Rh, root hair. Please click here to view a larger version of this figure.
