קבוע דיסוציאציה K הוא פרמטר מפתח כדי לתאר את זיקתו של קולטן (R) עבור שלה ליגנד (L). בהתבסס על חוק המסות, K מוגדר עבור האיזון, שבה RL מורכבים קולטן-ליגנד dissociates לתוך הקולטן R של ליגנד l:
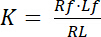 משוואה 1
משוואה 1
עם מדדי f המציין את מצב חינם/לא מאוגדים של קולטן וליגנד. הריכוז של המתחם קולטן-ליגנד, RL, זהה לריכוז ליגנד לקולטן מכורך Lb כמו ריכוז מוחלט של קולטן Rt הוא הסכום של קולטן חינם Rf וליגנד מכורך קולטן Rb = Lb, קבוע דיסוציאציה יכולה להיכתב גם בתור:
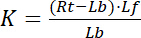 משוואה 2
משוואה 2
לכן, הרוויה תשואות Y, הגדיר את השבר של ליגנד מאוגד Lb ביחס של ריכוז מוחלט של קולטן Rt,
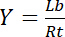 משוואה 3
משוואה 3
תלוי הריכוז של ליגנד חופשי Lf:
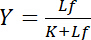 משוואה 4
משוואה 4
יחס היפרבולי זה מתאר עקומת קשירה של אינטראקציה קולטן-ליגנד ומראה העלילה שלו את הריכוז של ליגנד מאוגד Lb כפונקציה של הריכוז של ליגנד חופשי Lf. מן העקומה מחייב, ניתן לגזור את קבוע דיסוציאציה K כמו הריכוז של ליגנד חופשי-חצי-מקסימלי רוויה התשואה. יתר על כן, אלגוריתמים שונים כדי linearize מחייב עקומות הוקמו, כגון העלילה הכפולה-הדדיים מאת קלוץ1,2, או שינויי צורה בהתאם Scatchard או היינס (נבדקה ע י Bisswanger3). עם זאת, כל האלגוריתמים סובלים הבעיה שבה הערך המרבי של התשואה רוויה, אשר שקוורקים ניגשה בריכוזים גבוהים של ליגנד חופשי בתוך עקומת קשירה, חייב להיות מוערך בהערכה טרום גרפי, לכן הוא מועדת לטעויות.
בנוסף, הקביעה של עקומת קשירה דורשת כימות של ליגנד בחינם, מאוגד במהלך האיזון מחייב. למטרה זו, ליגנד חופשי צריך להיות הופרדו של ליגנד לקולטן מכורך לכמת. לכן, ליגנד לקולטן יש נבדלים בתכונותיהם, כגון ליגנד-חלבון לעומת חלבון קולטן. אם שני בני הזוג מחייב חלבונים, הם חייבים להיות להבדיל לגודלן, חיובים או תכונות אחרות מולקולרית. למרות זאת, כימות של ליגנד ריכוזים ב גישות איגוד בקנה מידה קטן הוא משימה קשה. תיוג רדיואקטיבי של ליגנד לעיתים קרובות כבר צורך לזהות את הריכוז נמוך של ליגנד מאוגד, במיוחד אם כמויות ניכרות של קולטנים לא היו זמינות או במחירים סבירים. יתר על כן, ליגנד לקולטן מכורך ייתכן מביצועם במהלך ואחרי בידוד באופן שעצירת. לפיכך, שיטות מורכבות, כגון שיווי משקל ג’ל סינון4, אלקטרופורזה נימי5הדופק proteolysis6, נדרשים לכמת ליגנד לקולטן מכורך ולהפריד אותו מן ליגנד חופשי.
בניגוד אלה מבחני הכריכה, ניסויים טיטור אינם דורשים ההפרדה כמותית של ליגנד כבול וחופשי. למטרה זו, הוא קולטן על ריכוז קבוע של טיטרציה עם ריכוזים שונים של ליגנד נוסף. באמצעות קשירה לקולטן, ליגנד מאוגד כוללת מאפיין biophysical אשר מבדיל ליגנד חופשי היא מדידה באמצעות, למשל, photometry, fluorometry או זיהוי נוגדנים. כך, האות S, אשר הוא יחסי הרוויה תשואות Y, וכתוצאה מכך גם לריכוז ליגנד לקולטן מכורך (Lb), ומתגלה כפונקציה של ריכוז מוחלט של ליגנד נוסף (Lt). פרמטרים, האות S וגם ריכוז מוחלט של ליגנד נוסף הם לכמת באופן ישיר וקל יותר מאשר הריכוזים של ליגנד כבול וחופשי. במיוחד, זיהוי הקולטן מכורך ליגנד מאת מקושרים-אנזים immunosorbent assay (אליסה) מותרות ההפחתה של אמצעי אחסון מדגם אל מתחת 100 µL וכן מדידות במקביל של מספר ריכוזים ליגנד צלחות רב טוב microtiter. ב טיטור אליסה, קולטן פיזית הספוחה על צלחת microtiter-ריכוז זהה, טיטרציה עם ליגנד מסיסים. הקולטן הוא מתאושש השטח פלסטיק בעיקרו של דבר על ידי ספיחה הידרופובי. ריכוז משטח של קולטן קיבוע בקורלציה עם הריכוז ציפוי של הקולטן ביחס לא לינארית, סביר על-פי Langmuir´s ספיחה איזותרמה7. בנוסף שהמספר הכולל של קולטן הספוחה מולקולות, מצב הפעילות שלהם היא פרמטר חשוב נוסף עבור מבחני טיטור. רק ללא יכולת תנועה קולטנים אשר שמרו ליגנד מחייב פעילות, הרלוונטיים עבור טיטור וזמינותו, בסופו של דבר לתרום ריכוז מוחלט של קולטנים הפעיל Rt של וזמינותו טיטור, אשר לא ניתן לקבוע באופן ישיר.
אתרים על פני השטח פלסטיק, אשר לא מכוסים על ידי רצפטור קיבוע נוטים לספוח חלבונים אחרים, כגון ליגנד. ספיחה פיסיקלית של ליגנד לאתרים משטח פלסטיק כזה יגרום ב אות דומה כמו ליגנד לקולטן מאוגד, עדיין באופן ספציפי. כדי להפחית את האות לא ספציפית, האתרים משטח פלסטיק של הלוחות microtiter אשר לא מצופה עם חלבון עדיין ייחסמו עם אלבומין שור (BSA). עם זאת, עבור כמה מבחני טיטור קולטן-ליגנד, רקע ספציפי אותות עלולים להיות שנצפו. ואז, סוכנים אחרים, חסימה, כגון פתרון של 0.2% ג’לטין או של 0.04% Tween 20, מומלץ.
לאחר הכריכה לקולטן ליגנד חופשי יוסר על ידי שני צעדים כביסה. ליגנד המאוגד נשארת עם הקולטן, שהוא מתאושש השטח פלסטיק של הבאר microtiter, לחלופין מחוזק על ידי קיבוע כימי. עבור העוקבים קוולנטיות הצלב הצמדה של ליגנד מאוגד, קולטן קיבוע עם גלוטראלדהיד, מוחלף החומר מאגר טריס עבור HEPES, ללא כל שינוי מחייב ליגנד. HEPES, בניגוד טריס, לא הפוך ללא פעיל גלוטראלדהיד. הצמדה הצלב קוולנטיות עם גלוטראלדהיד פותר את ליגנד מאוגד עם הקולטן שלו ומונע שלה דיסוציאציה במהלך הכביסה, דגירה והשלבים. לפיכך, האינטראקציה קולטן-ליגנד מוקפא מבחינה כימית, צווי עיקול טיטור אשר מושפע והשלבים של כביסה, דגירה. אולם, גלוטראלדהיד קיבוע עשוי כימית לשנות את ליגנד לקולטן באופן כזה כי האינטראקציה שלהם מצומצמת או ביטל. יתר על כן, שינוי של epitopes בתוך ליגנד עשוי להשתנות על זיקה מחייבת של הנוגדן גילוי, במיוחד אם נוגדן חד שבטי משמש לכמת ליגנד מאוגד. למרות וגם של אלה תופעות לוואי של קיבעון גלוטראלדהיד מתרחשת ב טיטור זו אליסה, הרגישות של הבדיקה לכיוון גלוטראלדהיד חייב להיבדק על כל אינטראקציה קולטן-ליגנד לפני הניסוי טיטור. לאחר קיבוע, גלוטראלדהיד עודף מוסר בשלושה צעדים כביסה עם מאגר המכיל טריס. טריס חלבונית קבוצות אלדהיד הנותרים, אשר עשוי להגיב nonspecifically עם זיהוי נוגדנים בשלב שלאחר מכן.
הכמות של ליגנד מאוגד לכמתו מקושרים-אנזים נוגדנים, המספקים אות אליסה בהשוואת ס זה מותווים לעומת ה ליגנד הכולל ריכוז L-t הוסיף את כל טוב. למרות הרכישה לקל יותר, העקומה טיטור אינה פונקציה היפרבולית בניגוד העקומה מחייב. יתר על כן, זה כבר לא ברור כיצד לחשב את קבוע דיסוציאציה K מ עיקול טיטור. למרות אלגוריתמים כדי linearize spectroscopically רכשה טיטור עקומות דווחו באופן עצמאי על-ידי Stockell8 ו- היי, Weischet9, הם התמוטטו עקב חוסר הבהירות של הערכת האות המרבי ערך התשואה רוויה ניגש בריכוזים גבוהים של ליגנד נוסף.
כאן, טיטור אליסה, באלגוריתם רגרסיה ליניארי מתוארים להפיק את קבוע דיסוציאציה K לאינטראקציה ליגנד קולטן של עיקול טיטור. פרוטוקול זה הוא דוגמה עבור האינטראקציה של קולגן מחייב A-התחום של α2β1 אינטגרין עם מעכב נגזר ארס הנחש. אינטגרינים הם מולקולות אדהזיה התא, אשר לתווך המעגנה תאים של מטריצה חוץ-תאית שמסביב או כבסיס קרום המרתף10,11. יתר על כן, אינטגרינים להעביר חשוב אותות בין תאים של מטריצה חוץ-תאית על-ידי גיוס נוספים מולקולות איתות ויוצרים האברונים חדש, adhesomes, עם תא-מטריצת אינטראקציה12,13, 14. קולגן, ליגנד של אינטגרין α2β1, הוא החלבון הנפוץ ביותר בגוף האדם, הוא מרכיב קריטי פיגומים של רקמת חיבור ה-15. האינטראקציה בין α2β1 אינטגרין קולגן מתווך על ידי A-התחום של יחידת משנה α2 אינטגרין. אינטגרין α2A-התחום מכיל הקטיון כלט, אשר נדרש עבור איגוד קולגן ומייצב את המבנה שלה. טופס פראי-סוג, כמו גם מוטציות של מתחם α2A, כמו זו שבה שאריות השטח החשופים Y216 הוחלף עבור גליצין, בקלות ניתן המיוצר recombinantly במערכת הביטוי חיידקי, מבודדים באמצעות שלהם oligo-שלו-תגיות עם נינטה superflow טורים דיאליזה הבאים נגד באגירה טריס תמיסת מלח (TBS; 50 מ”מ טריס/HCl, pH 7.4, 150 מ מ NaCl) המכילה 2 מ מ MgCl216. ריכוזים שלהם היו נחושים עם וזמינותו חומצה bicinchoninic (BCA), purities שלהם נבדק על ידי מרחביות קונבנציונלי-דף, מוכתם R250 כחול Coomassie-מבריק.
האינטראקציה בין α2β1 אינטגרין קולגן חסומה על-ידי איגוד של הרכיב ארס הנחש, rhodocetin, מ ה16,בור צפע (Calloselasma rhodostoma)17. משמש של ליגנד מסיס ב טיטור זו אליסה, rhodocetin היה טהור מן גולמי ארס כפי שתואר לעיל16. זה הוא מומס באגירה HEPES תמיסת מלח (HBS; 10 מ מ HEPES/NaOH, pH 7.4, 150 מ מ NaCl) ומאוחסן קפוא ב-20 ° C. הריכוז שלו נקבע על ידי BCA, הטוהר שלו הוכח על ידי עמודים מרחביות. כמו אנטגוניסט, rhodocetin לא רק חוסם קולגן איגוד אינטגרין α2β1 A-מחשבים, אבל גם מייצבת את קונפורמציה לא פעיל אינטגרין ובכך מונע כל איתות של קולגן לתוך תאים או טסיות18. זה חשיבות הביו-רפואית לקבוע את קבוע דיסוציאציה של rhodocetin עם המטרה הקולטן שלו, ובכך לפענח את המנגנון המולקולרי ואת תרופות פוטנציאליות למשל, כסוכן בשבץ antithrombotic. למטרה זו, טיטור אליסה מתואר כולל את ההערכה שלו, אשר חל כמעט כל קולטן-ליגנד אינטראקציה עם סטויכיומטריה אינטראקציה עם 1:1.
