Figure 1 indicates the time required for three-dimensional germline analysis. L4 hermaphrodites incubated at 20 °C were dissected to isolate germlines and stained with DAPI, phalloidin, and antibodies against germline proteins. Germlines are imaged using confocal microscopy. Staining and confocal microscopy requires approximately 24 h. Computational analysis for the complete germline requires 10 – 15 min to count the number and position of nuclei, identify the protein distribution, analyze the cytoskeletal structure and score the number of sperm. Complete manual analysis requires over 2 h per germline and is subject to operator influence, therefore, our automated high-throughput method reduces the analysis time and enhances the reproducibility.
Automated nuclei scoring reveals that a germline arm houses approximately 1,000 – 1,200 nuclei (Figure 2C), corresponding well with previously published manual scoring data1. To confirm the accuracy of scoring, multiple mutants and different conditions are used (Figure 2D-G). For instance, we compared rnp-8(tm435), cpb-3(bt17), and glp-1(e2141) mutant worms to wild type animals. Automated counting reproduced the known reduction of number of nuclei in cpb-3 and glp-1 mutant germlines14,15,16,17. A severe reduction in nuclei number was also observed in the glp-1(e2141) temperature-sensitive mutant when incubated at 25 °C. The distribution of nuclei is dependent on the stage of differentiation. The mitotic region, which contains approximately 250 nuclei in wild type, is tightly-packed with nuclei compared to the rest of the germline (Figure 3)1. The spacing between mitotic nuclei is minimal and nuclei are located throughout the mitotic region. However, as the nuclei move away from distal end, they appear more towards the circumference of the germline (Figure 3). The change in distribution starts at the transition zone and completes as the nuclei enter meiosis/pachytene.
The germline has specific cytoskeletal structures at different stages of differentiation. We visualized F-actin by staining the germline with phalloidin. The distribution of F-actin from the distal to proximal end of the germline was distinct at each region (Figure 3 and Figure 5). In general, there are two separate structures that appear as a 'cylinder within cylinder.' At the mitotic region, inner actin appears as a solid mass with a specific shape (Figure 3). As the germline reaches the transition zone, the inner actin assumes a more cylindrical structure and it becomes a hollow cylinder as it reaches late pachytene. The second layer of actin covers the inner layer giving shape to the germline. This 'cylinder within cylinder' actin structure disappears towards the oocyte region (Figure 5). At the oocyte region, actin appears as thick fibers. The oocytes are separated by an actin rachis, though it appears to be dynamic to allow the movement of oocytes to the spermatheca. Finally, actin in the spermatheca also forms thick bundles of actin fibers. However, the fibers appear to be packed closer than in the oocyte region. The cytoskeletal structure appears to complement the distribution patterns of nuclei (Figure 3). At the mitotic region, the nuclei appear to be placed around the inner actin structure. As they reach meiosis/pachytene, the nuclei organize between the two actin cylinders leaving the middle of the germline mostly devoid of nuclei (Figure 3). This could probably aid uninterrupted cytoplasmic streaming in the germline. Germlines were stained with an antibody against the REC-8 protein and analyzed by confocal microscopy. REC-8 is homogeneously distributed around early nuclei of the germline and is later cleaved and degraded during meiosis18. REC-8 distribution in three-dimensional cross-sectional analysis of the germline shows the distribution of protein in the mitotic region (Figure 4).
Three-dimensional rendering of the proximal end of the germline includes the spermatheca and first three oocytes distal to the spermatheca. Our analysis recognized an average of 151 sperm in each spermatheca (n = 18). This corresponds to the published literature, indicating the reliability of the method (Figure 5F)19. This analysis may be performed together with cytoskeletal or protein distribution studies. The C. elegans genome is distributed among six chromosomes. In fully developed wild type oocytes, these chromosomes are visible and can be counted. This chromosome separation can be used to study chromosome stability or other defects associated with oocyte development. By three-dimensional rendering, the number of chromosomes can be visualized, and if the chromosomes are separated properly, the number can be accurately obtained (Figure 5).
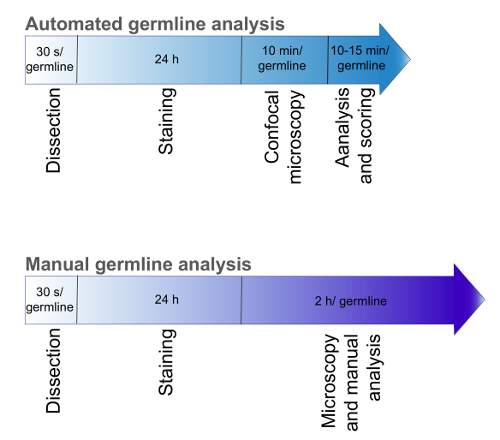
Figure 1. Timeline for automated analysis. Time comparison between manual and computational analysis of the C. elegans germline. Please click here to view a larger version of this figure.
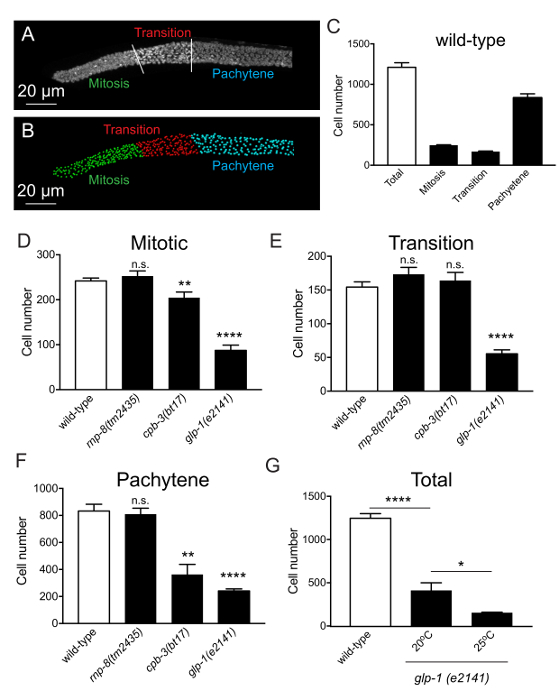
Figure 2. Nuclei distribution in the germline. (A) DAPI stained germline showing mitotic, transition, and pachytene/meiotic regions. (B) Computational three-dimensional model of the germline. (C) Scoring for the total number of nuclei in wild type germlines. (D-F) Scoring for the number of nuclei at mitotic, transition, and meiotic regions of wild type, rnp-8, cpb-3, and glp-1 mutant germlines. (G) Comparison in the total number of nuclei in the glp-1 mutant at 20 °C and 25 °C. Error bar represents standard error. Student's test. ****n <0.0001, ***n <0.001, **n <0.01, *n <0.05. Please click here to view a larger version of this figure.
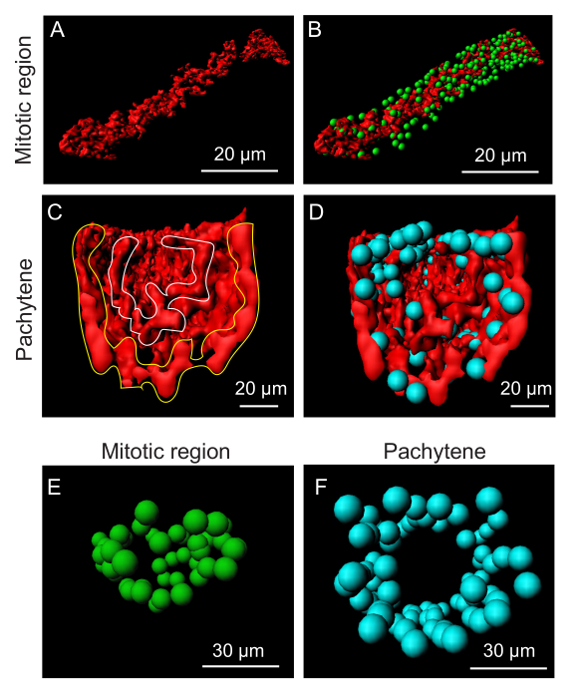
Figure 3. Cytoskeletal structure of the germline. (A-B) Organization of the inner actin (red) cytoskeleton of the germline at the mitotic region. The inner actin has a diffuse structure compared to outer actin at the mitotic region. (C-D) At the pachytene region, the cytoskeleton assumes a cylindrical structure by forming two cylinders between which the nuclei (blue) are organized. (E-F) Organization of germline nuclei in the mitotic and pachytene regions. The distribution of nuclei is more towards the circumference of the 'germtube' at the pachytene region compared to mitotic region (green) and middle of the germline become devoid of nuclei. Please click here to view a larger version of this figure.
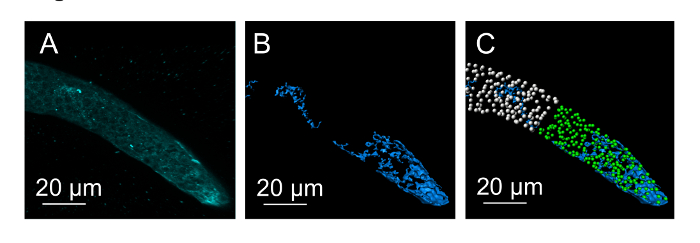
Figure 4. REC-8 expression in the mitotic region. (A) Micrograph showing the distal end of a REC-8-stained germline. (B-C) Three-dimensional rendering of REC-8 staining (shown in blue) and distribution of protein between the mitotic nuclei (green). REC-8 staining is less abundant proximal to the mitotic region (marked by white dots representing nuclei). Please click here to view a larger version of this figure.
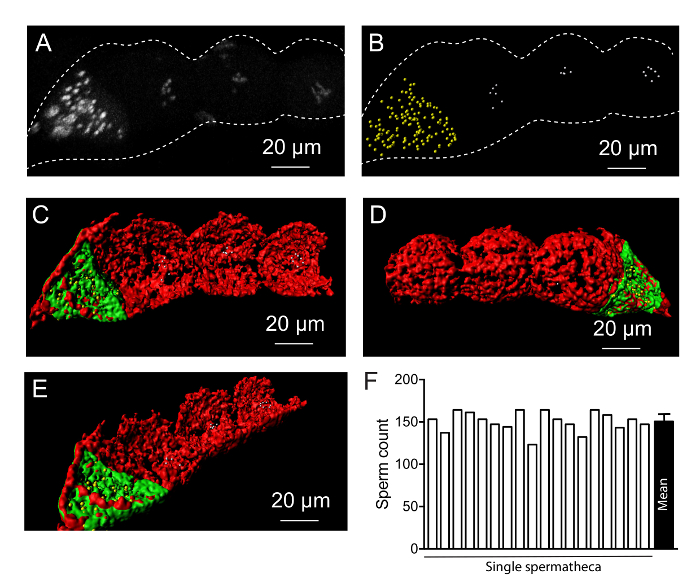
Figure 5. Germline structure at the proximal end. (A) DAPI staining of the germline at the proximal end. (B) Three-dimensional rendering of DAPI staining showing sperm in the spermatheca (yellow) and chromosomes in the oocytes (white). (C-E) Three-dimensional cytoskeletal structure at the proximal end of the germline. Red marks the complete proximal end and green marks the spermatheca. Cross sectional analysis shows that the cytoskeleton not only forms a filamentous structure of actin around oocytes, but also separates between oocytes. (F) Graph showing the number of sperm in each spermatheca and average obtained from multiple spermatheca (n = 18). Error bar represents standard error. Please click here to view a larger version of this figure.
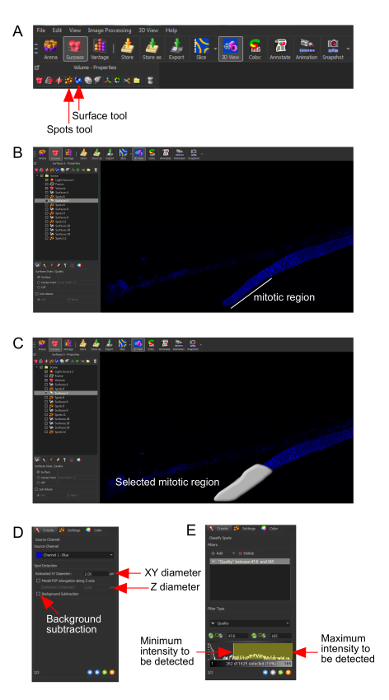
Supplementary Figure 1. Software tools description. (A) The spots and surface tools for detecting nuclei and protein staining. (B-C) The selection of three-dimensional region of interest using the surface tool. (D) Defining the XY-diameter, using spots tool, for nuclei detection. Depending on the region of interest and staining, the Z-axis diameter and background correction can be used. (E) Minimum and maximum intensity threshold detection. A similar approach can be used for surface function where instead of defining the diameter, and the surface details can be defined. Please click here to view a larger version of this figure.
