العديد من العمليات البيولوجية التي تحدث في مواقع التفاعلات خلية خلية، مثل خلية خلية التصاق1،2،3، خلية خلية الانصهار4 والاعتراف الخلوية5. هذه الأحداث تمثل أهمية خاصة أثناء تطوير الكائنات الحية متعددة الخلايا، وخلية خلية الاتصالات، مثلاً، أثناء الاستجابات المناعية. وساطة هذه العمليات عادة من البروتينات التي تكون مترجمة على السطح، أي في غشاء البلازما (م) من الخلايا المجاورة وتخضع لتفاعلات معينة في خلية خلية الاتصال التي هي على وجه التحديد الخاضعة للتنظيم في المكان والزمان. في كثير من الحالات، هذه التفاعلات المباشرة هومو-أو التفاعلات عبر الغيروية البروتين-بروتين، ولكن قد ينطوي أيضا على أيونات أو يغاندس بوصفها linkers خارج الخلية1. على الرغم من أهمية أساسية، وهناك نقص في فحوصات سبر هذه تفاعلات البروتين البروتين محددة مباشرة في البيئة الأصلية للخلايا الحية. تتطلب العديد من الأساليب أما تعطيل الخلية (مثل فحوصات الكيمياء الحيوية مثل شركة إيمونوبريسيبيتيشن6)، التثبيت (مثلاً، بعض تقنيات بصرية مجهرية فائقة القرار والمجهر الإلكتروني لخلية خلية اتصالات7)، أو هي غير محددة، مثل التجميع/التصاق فحوصات8،9. للتغلب على هذه المشكلة، تم تنفيذ fluorescence تقنيات استناداً إلى الأسفار الرنين الطاقة نقل (الحنق)10 أو الأسفار التكامل11. ومع ذلك، تتطلب هذه الطرق تحقيقا لمسافات صغيرة بما فيه الكفاية بين فلوروفوريس، تسميات الفلورسنت على الجانب خارج الخلية البروتينات10، يحتمل أن تتداخل مع التفاعلات عبر .
نقدم هنا، تحليل القائم على الأسفار بديلة لتفاعلات البروتين البروتين في الخلية جهات الاتصال. هذا النهج يجمع بين النهج عبر الارتباط الأسفار (الأسفار المسح الضوئي عبر ارتباط مطيافية (سفككس)، وعدد عبر الارتباط والسطوع (ccN & ب)) وخلط الخلايا معربا عن بناء انصهار من البروتين من الفائدة، مثلاً، مستقبلات التصاق. تتم تسمية مستقبلات التحقيق في الخليتين التفاعل مع البروتينات الفلورية طيفيا فصل اثنين (FPs)، من داخل الخلايا الجانب (انظر الشكل 1A).
أساليب العاملين تقوم على التحليل الإحصائي للأسفار التقلبات الناجمة عن الحركة انتشارية البروتينات الفلورية الانصهار من خلال حجم ليزر [كنفوكل] مجهر مسح التنسيق. أكثر في التفاصيل، ويَسْبِر المقايسة نشر المشارك من البروتينات ذات الاهتمام في كلا PMs المجاورة في خلية خلية الاتصالات. إذا البروتينات الخضوع عبر التفاعلات، سيحمل هذه المجمعات عبر البروتينات الفلورية التي تنبعث منها في كل القنوات الطيفية، تسبب تقلبات fluorescence مرتبطة من بواعث كلا. من ناحية أخرى، في حالة حدوث لا ملزمة، تقلبات عدد من البروتينات التي تواجه الدورة الشهرية ستكون مستقلة، مما تسبب في لا تقلبات مرتبطة. اكتساب يمكن أن يؤديها بطريقتين: 1) سفككس يستند على شكل خط المسح الضوئي عبر الاتصال خلية خلية ويَسْبِر فعالية التفاعلات في بقعة تقع في منطقة الاتصال. من خلال تحليل الزمانية لتقلبات الأسفار، سفككس يوفر أيضا معلومات حيوية، أي معاملات نشر مجمعات البروتين؛ 2) ccN & ب يستند إلى إجراء تحليل بيكسيلويسي لسلسلة من الصور التي حصلت في مناطق الاتصال خلية خلية. قد القدرة على التحقيق وخريطة التفاعلات على طول كامل الاتصال بالمنطقة (في المستوى البؤري واحد)، ولكن لا يقدم معلومات عن ديناميات. يمكن الجمع بين كلا الأسلوبين مع تحليل للسطوع الجزيئية، أي إشارة fluorescence متوسط المنبعثة في الوحدة الزمنية من مجمعات البروتين نشرها مفردة، وهكذا، تقديم تقديرات ل stoichiometry مجمعات البروتين في خلية الاتصالات.
في هذه المقالة، نحن نقدم بروتوكولات مفصلة لإعداد عينة وأداة المعايرة، الحصول على البيانات وتحليلها لإجراء التحليل المقدم على ليزر [كنفوكل] تجارية المسح مجهر. يمكن إجراء هذه التجارب على أي صك مجهزة بالعد فوتون أو كاشفات التناظرية وموضوعي مع ارتفاع الفتحة العددية. علينا مواصلة مناقشة الخطوات الحاسمة للبروتوكول وتقديم مخططات تصحيح للعديد من العمليات التي تسبب تقلبات إشارة أرتيفاكتوال، مثلاً، للكشف عن الضوضاء، وحركة فوتوبليتشينج أو الخلية. وضعت أصلاً للتحقيق في التفاعلات بين الخلايا ملتصقة، التحليل يمكن تعديلها لتعليق الخلايا، أو تكييفها لنظم الغشاء النموذجي، مثلاً، أونيلاميلار العملاقة حويصلات (جوفس) أو البلازما العملاقة غشاء الحويصلات (جبمفس)، مما يسمح التحديد الكمي للتفاعل في البيئات المختلفة الدهن، أو في حالة عدم12،سيتوسكيليتون المنظمة13.
المسح الضوئي الطيفي الارتباط عبر الأسفار هو نسخة معدلة من الأسفار عبر ارتباط مطيافية14 وتم تصميمه خصيصا للتحقيق في ديناميات انتشارية بطيئة في الأغشية الدهنية15. أنه يستند إلى عملية شراء مسح خط عمودي إلى الساعة التي تحتوي على بروتينات الفلورسنت للفائدة. للتحقيق في التفاعلات بين نوعين من أنواع البروتين المسمى بشكل مختلف، يتم اكتساب في قناتين طيفية استخدام اثنين من خطوط الليزر وهما كشف windows ل fluorophores طيفيا المنفصلين عن ذويهم. سبب ديناميات نشر بطيئة من البروتينات في الساعة (D≤ ~ 1/ق2ميكرومتر)، يمكن أن يؤديها قياس خالية من الحديث عبر مخطط الإثارة من سطر إلى سطر15بالتناوب. يبدأ التحليل: 1) محاذاة خوارزمية تصحيح لحركة الخلية الجانبية استناداً إلى بلوكويسي في المتوسط من خطوط ~ 1000، 2) تحديد الموقف مع الموقف، أي الساعة fluorescence أقصى إشارة، في كل كتلة و 3) تحويل جميع القطع لمشترك منشأ12،15، كل على حدة في كل قناة. ثم، يتم تحديد تلقائي لتناظر الساعة بكسل بتحديد المنطقة الوسطى من نوبة ضبابي من مجموع كافة الأسطر الانحياز (أي، مركز ± 2.5σ). تكامل الإشارة في كل سطر غلة السلسلة الزمنية fluorescence غشاء F(t) في كل قناة (ز = الأخضر قناة، r = القناة الحمراء). علما بأن حجم بكسل يجب أن تكون صغيرة بما يكفي، مثلاً، < 200 نانومتر، إعادة بناء شكل النقطة نشر وظيفة والعثور على مركزها، المقابلة لموقف رئيس الوزراء. حضور فوتوبليتشينج كبيرة، قد يكون على غرار مع دالة آسيه مزدوجة السلسلة الزمنية الأسفار في كل قناة وثم تم تصحيحها بالصيغة التالية:16
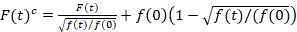 . (1)
. (1)
من المهم ملاحظة أن هذه الصيغة بفعالية تصحيح كلا من الاتساع ونشر الأوقات التي تم الحصول عليها من تحليل الارتباط F(t)ج، مقارنة بتقديرات المعلمة التي يمكن الحصول على من غير المصححة F(t). ثم، مهام الارتباط التلقائي والصليب (أكفس/أطر التعاون القطري) وتحسب للأسفار إشارات:
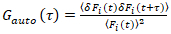 ، (2).
، (2).
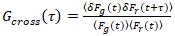 ، (3).
، (3).
حيث δوأنا = وأنا(t)- 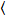 وأنا(t)
وأنا(t)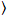 و أنا = ز، والبحث والتطوير.
و أنا = ز، والبحث والتطوير.
نموذج نشر ثنائي الأبعاد ثم مزودة بجميع مهام الارتباط (لجنة الأمن الغذائي العالمي):
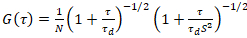 . (4)
. (4)
هنا، تشير N إلى عدد البروتينات الفلورية في حجم المراقبة و τد وقت نشرها لكل قناة. هذا النموذج يأخذ في الاعتبار أن في وصف الإعداد التجريبية، يحدث انتشار البروتينات في الساعة في الطائرة x-z، خلافا لتكوين علاقة الفلورية استخداماً التحليل الطيفي (FCS) تجارب على الأغشية السبر نشر في الطائرة x-y من حجم [كنفوكل]17. الخصر ث0 وعامل البنية S، واصفاً الإطالة wض حجم التنسيق في z, S = ثضث0، يتم الحصول عليها من قياس معايرة FCS نقطة أدوا بالأصباغ طيفيا مماثلة ونفس إعدادات البصرية استخدام القيم الموجودة بالفعل لأن معامل نشر دصبغ:
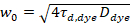 ، (5).
، (5).
حيث τد، وصبغ هو وقت نشر متوسط قياس جزيئات الصبغة، التي تم الحصول عليها من تركيب نموذج لنشر ثلاثية الأبعاد للبيانات، ومراعاة التحولات حساب الكسر ر كل ن الجزيئات إلى الدولة الثلاثي مع ثابت وقت ττ:
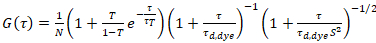 . (6)
. (6)
وأخيراً، نشر معاملات (د) وقيم السطوع الجزيئية (اليورو) والصليب-الارتباط النسبي للبيانات سفككس (rel.cc.) يتم حساب كما يلي:
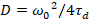 ، (7).
، (7).
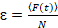 ، (8).
، (8).
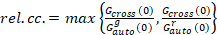 ، (9).
، (9).
زالصليب(0) من حيث السعة للدالة عبر الارتباط و 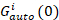 هو السعة للدالة ترابط تلقائي في القناة-ال أنا.
هو السعة للدالة ترابط تلقائي في القناة-ال أنا.
هذا التعريف نسبي الصليب-العلاقة المتبادلة، أي استخدام كحد أقصى بدلاً من يعني في المعادلة 9، يأخذ في الاعتبار أن يقتصر الحد الأقصى لعدد من المجمعات من نوعين من أنواع البروتين الحالية بتركيزات مختلفة الأنواع الموجودة في عدد أقل.
عدد عبر الارتباط وسطوع يستند إلى تحليل هذه لحظة لشدة الأسفار لكل بكسل من رصة صور اكتسبت على مر الزمن في موقع ثابت في العينة، تتألف عادة من ~ 100-200 الإطارات، مع اثنين الطيفية القنوات ( g = أخضر القناة، r = القناة الحمراء). من الوسط الزماني أناأنا والفرق 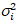 ، وتحسب سطوع الجزيئية اليوروi وعدد نأنا في كل بكسل وقناة الأطياف (أنا = g, r)18:
، وتحسب سطوع الجزيئية اليوروi وعدد نأنا في كل بكسل وقناة الأطياف (أنا = g, r)18:
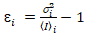 ، (10).
، (10).
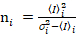 . (11)
. (11)
من المهم ملاحظة أن معادلات معينة تنطبق على الحالة المثالية لكاشف فوتون العد الحقيقي. لأنظمة الكشف التناظرية، تطبيق المعادلات التالية19،20:
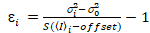 ، (12).
، (12).
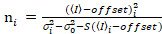 . (13)
. (13)
هنا، هو S معامل التحويل بين الفوتونات المكتشفة والتهم الرقمية المسجلة، 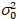 ضجيج قراءات و إزاحة يشير إلى الكشف عن كثافة الإزاحة. عموما، يجب معايرة هذه الكميات، لأي نوع الكاشف، استناداً إلى قياس الفرق كاشف كدالة كثافة لإنارة مطرد19، مثلاً، سطح معدني عاكس أو حل صبغ المجففة. يمكن تحديد الإزاحة عن طريق قياس معدل العد في عينة دون ضوء الإثارة. عن طريق إجراء انحدار خطي للفرق المرتبطة بالكشف عن
ضجيج قراءات و إزاحة يشير إلى الكشف عن كثافة الإزاحة. عموما، يجب معايرة هذه الكميات، لأي نوع الكاشف، استناداً إلى قياس الفرق كاشف كدالة كثافة لإنارة مطرد19، مثلاً، سطح معدني عاكس أو حل صبغ المجففة. يمكن تحديد الإزاحة عن طريق قياس معدل العد في عينة دون ضوء الإثارة. عن طريق إجراء انحدار خطي للفرق المرتبطة بالكشف عن 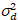 مقابل الأرض كثافة (أنا), S و يمكن أن تكون حازمة19:
مقابل الأرض كثافة (أنا), S و يمكن أن تكون حازمة19:
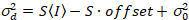 . (14)
. (14)
وأخيراً، يحسب في كل بكسل سطوع عبر الارتباط ويتم تعريفها بشكل عام21
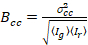 ، (15).
، (15).
حيث 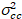 هو الصليب-الفرق
هو الصليب-الفرق 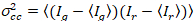 .
.
من أجل تصفية تقلبات المعمرة، تتم جميع ccN & حسابات ب بعد النقل التصفية، بشكل مستقل لكل بكسل22. بإيجاز، ni, اليوروأنا (أنا = g, r) وتحسب بcc في انزلاق قطاعات مثل إطارات 8-15. يمكن أن متوسط القيم التي تم الحصول عليها ومن ثم ثم الحصول على بكسل النهائي القيم عدد والسطوع.
تحليل ستويتشيوميتري
بغية تقدير stoichiometry مجمعات البروتين في الخلية جهات الاتصال، يمكن تحليل سطوع الجزيئية بشكل منفصل في كل قناة طيفية سفككس أو ccN & ب البيانات. في سفككس، يتم الحصول على قيمة سطوع واحدة كل قياس في كل قناة. في ccN & B، يتم الحصول على الرسم بياني سطوع لكل بكسل المقابلة لجهة الاتصال خلية خلية وقيمة متوسط (أو الوسيط) يمكن استخدامها سطوع الممثل للقياس. عن طريق إجراء نفس التحليل على مرجع أحادي، يمكن تطبيع جميع قيم السطوع مباشرة الحصول على الدولة oligomeric متوسط مجمعات البروتين تم الكشف عنها. في هذه المرحلة، من المهم تصحيح لوجود FPs غير الفلورية قد يؤدي التقليل من الدولة أوليجوميريك. وهذا عادة ما يقوم بقياس سطوع23،البروتين مرجع هومو dimeric24 استخدام سفكس لون واحد أو عدد والسطوع (ن وب).
