Многие биологические процессы происходят на объектах ячеек взаимодействий, например, ячеек адгезии1,2,3, слияние ячеек4 и клеточных признание5. Такие мероприятия особенно важны во время развития многоклеточных организмов и для связи ячеек, например, во время иммунных реакций. Эти процессы обычно опосредовано белки, которые локализованы на поверхности, то есть, на плазматической мембраны (PM) соседних клеток и проходят специфических взаимодействий в контакт ячеек, которые точно регулируется в пространстве и времени. Во многих случаях эти взаимодействия прямого гомо – или гетеротипичной белок белковых взаимодействий транс , но может также включать ионов или лигандов, действуя как внеклеточного линкеры1. Несмотря на основополагающее значение, отсутствует анализов зондирующего эти конкретные белок белковых взаимодействий непосредственно в родной среде живых клеток. Многие методы либо требуют разрушения клеток (например, биохимических анализов как co иммунопреципитация6), фиксация (например, некоторые методы оптической микроскопии суперразрешением и электронной микроскопии ячеек Контакты7), или являются неспецифическими, например, агрегации / сцепления assays8,9. Для преодоления этой проблемы, были выполнены флуоресценции методы, основанные на флуоресценции резонансные энергии передачи (ЛАДА)10 или флуоресценции дополнения11. Однако чтобы добиться достаточно небольших расстояниях между флуорофоров, эти методы требуют флуоресцентные метки на стороне внеклеточных белков10, потенциально мешать транс взаимодействий.
Здесь мы представляем альтернативной основе флуоресценции assay для взаимодействий протеин протеина в ячейке контактов. Этот подход сочетает в себе флуоресценции кросс корреляции подходы (сканирование флуоресценции кросс корреляции спектроскопии (sFCCS), кросс корреляции номер и яркость (ccN & B)) и смешивания клеток, выражая конструкцию синтеза белка интерес, например, рецептор адгезии. Исследованы рецепторов в двух взаимодействующих клеток помечены с двумя спектрально разлученных флуоресцентных белков (FPs), от внутриклеточного стороне (см. рис. 1A).
Занятых методы основаны на статистическом анализе флуоресценции колебаний, вызванных диффузионного движения флуоресцентные синтез белков через фокуса тома конфокальный лазерный сканирующий микроскоп. Более подробно, assay зонды совместное распространение протеинов интереса в обеих соседних PMs в ячейке контактов. Если белки проходят транс взаимодействий, эти комплексы транс будет нести флуоресцентных белков в обоих спектральных каналов, вызывая колебания коррелированных флуоресценции обоих излучателей. С другой стороны если привязка не происходит, число колебаний белков сталкивается ПМС будет независимым, вызывая не коррелированные колебания. Приобретение может быть выполнена двумя способами: 1) sFCCS основана на линии образный сканирования через контакт клеток и эффективно зонды взаимодействий в месте, расположенная в регионе контакта. Через временной анализ колебаний флуоресценции sFCCS обеспечивает также динамика информацию, т.е. коэффициенты диффузии белковых комплексов; 2) ccN b & B на основе pixel-wise анализа последовательности изображений, приобретенных в регионах контактных ячеек. Он имеет возможность зонда и карта взаимодействия вдоль всего связаться региона (в одной фокальной плоскости), но не предоставляет информацию о динамике. Оба метода могут быть объединены с анализ молекулярных яркость, т.е. средняя флуоресценции сигнал излучаемый одного диффундирующих белковых комплексов в единицу времени и, таким образом, дают оценки стехиометрии белковых комплексов в Контакты ячеек.
В этой статье мы предоставляем подробные протоколы для пробоподготовки, поверки, сбора и анализа данных для выполнения представленных пробу на коммерческих конфокальный лазерный сканирующий микроскоп. Эксперименты могут выполняться на любом инструменте, оснащенных Фотон подсчета или аналоговые датчики и объективным с высоким числовая апертура. Мы далее обсудить важнейшие шаги протокола и обеспечивают коррекцию схемы для нескольких процессов, вызывая колебания артефакта сигнала, например, детектор шум, Фотообесцвечивание или ячейки движения. Первоначально разработанный прозондирует область взаимодействия между адэрентных клеток, assay может быть изменена для подвески клетки, или адаптированы к модели мембранных систем, например, гигантский однослойных везикул (GUVs) или гигантский плазменный мембрана везикулы (GPMVs), что позволяет количественное определение взаимодействий в средах различных липидов или в отсутствие организованной цитоскелета12,13.
Сканирование на страницу отдела Флуоресценция кросс корреляции представляет собой модифицированную версию флуоресцентной спектроскопии кросс корреляции14 и была специально разработана для зонда медленно диффузионное динамика в липидных мембран15. Он основан на линии сканирования приобретение перпендикулярно вечера, содержащий флуоресцентные белки интерес. Прозондирует область взаимодействия двух видов по-разному помечены белка, приобретение осуществляется в двух спектральных каналов, с помощью двух лазерных линий и два окна обнаружения для спектрально разлученных флуорофоров. Из-за медленного распространения динамики белков в ТЧ (D≤ ~ 1 мкм2/s), кросс talk свободные измерения могут быть выполнены путем чередования схемы возбуждения от линии к линии15. Анализ начинается с: 1) выравнивания алгоритм исправления для боковых клеточного движения на основе block-wise в среднем ~ 1000 линий, 2) определение положения с максимальной флуоресценции сигналато есть, вечера позиции, в каждом блоке и 3) Ветра из всех блоков для общего происхождения12,15, отдельно в каждом канале. Затем автоматический выбор пикселов, соответствующих вечера выполняется, выбрав Центральный регион от Гаусса fit суммы все выровненные линии (т.е., центр ± 2.5σ). Интеграция сигнала в каждой строке дает мембраны флуоресценции рядов F(t) в каждом канале (g = зеленый канал, r = красный канал). Обратите внимание, что размер пикселя должна быть небольшой достаточно, например, < 200 Нм, чтобы восстановить форму точки распространения функция и найти свой центр, соответствующие положения пп. При наличии существенных Фотообесцвечивание, флуоресценции временных рядов в каждом канале может быть смоделированы с двойной экспоненциальная функция и затем исправлены с помощью следующей формулы:16
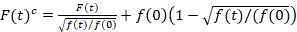 . (1)
. (1)
Важно отметить, что эта формула эффективно исправляет амплитуд и диффузии раз, полученные из корреляционного анализа F(t)c, по сравнению с оценки параметров, которые будут получены от нескорректированной F(t). Затем, функции auto – и кросс корреляции (ACFs / РСС) флуоресценции сигналы рассчитываются:
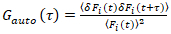 , (2).
, (2).
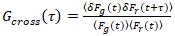 , (3).
, (3).
где δF,я = F,я(t) – 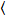 F,я(t)
F,я(t)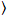 и я = g, r.
и я = g, r.
Двумерный диффузионная модель после этого приспособлена для всех корреляционных функций (CFs):
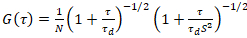 . (4)
. (4)
Здесь N обозначает количество флуоресцентных белков объема наблюдения и τd диффузии время для каждого канала. Эта модель принимает во внимание, что в параметре описанных экспериментальных, диффузии белков в Личку происходит в плоскости x-z, в отличие от часто используемые конфигурации флуоресценции корреляции спектроскопии (FCS) экспериментов на мембраны зондирование Диффузия в плоскости x-y конфокальный объем17. Талии w0 и структура фактора S, описывая удлинение wz фокуса тома в z, S = wz/w0, полученные из измерения Калибровка точки FCS, выступал с спектрально аналогичные красители и оптических параметров используя уже имеющиеся значения для коэффициента диффузии Dкраска:
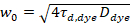 , (5).
, (5).
где τd, краска это измеренное среднее диффузии время краситель молекулы, полученные из установки модели для трехмерных распространения данных, принимая во внимание переходы дроби T всех N молекул в триплет государство с константой времени ττ:
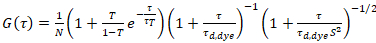 . (6)
. (6)
Наконец коэффициенты диффузии (D), значения молекулярных яркости (ε) и относительной кросс корреляции данных sFCCS (rel.cc.) рассчитываются следующим образом:
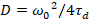 , (7).
, (7).
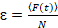 , (8).
, (8).
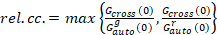 , (9).
, (9).
где Gкрест(0) является амплитуда функции кросс корреляции и 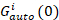 – амплитуда автокорреляционной функции в i-й канал.
– амплитуда автокорреляционной функции в i-й канал.
Это определение относительной кросс-корреляции, т.е. вместо Макс означает в уравнение 9, принимает во внимание, что максимальное количество комплексов двух видов белка в различных концентрациях ограничен в меньшее число видов.
Кросс корреляции номер и яркость основан на момент анализа интенсивности флуоресценции для каждого пикселя стека изображений приобрели со временем в фиксированном положении в образце, обычно состоящая из ~ 100-200 фоторамки, с двумя спектральных каналов () g = зеленый канал, r = красный канал). От височной среднего яя и дисперсией 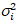 , молекулярный яркость εi и число n,я рассчитываются каждый пиксель и спектральных каналов (я = g, r)18:
, молекулярный яркость εi и число n,я рассчитываются каждый пиксель и спектральных каналов (я = g, r)18:
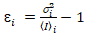 , (10).
, (10).
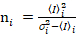 . (11)
. (11)
Важно отметить, что данного уравнения применяются в идеальном случае истинный Фотон подсчет детектора. Для обнаружения аналоговых систем следующие уравнения применяются19,20:
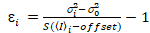 , (12).
, (12).
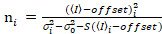 . (13)
. (13)
Здесь, S является коэффициент пересчета между обнаруженные фотоны и записи цифровых отсчетов, 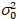 это шум считывания и смещение ссылается на смещение интенсивности детектор. Как правило эти количества должны калиброванные, для любого типа детектора, основанный на измерении детектор дисперсии в зависимости от интенсивности для постоянного освещения19, например, отражающей поверхности металла или сушеные краситель решение. Смещение может быть определена путем измерения скорость счета для образца без возбуждения света. Выполнив линейную регрессию детектор связанные дисперсии
это шум считывания и смещение ссылается на смещение интенсивности детектор. Как правило эти количества должны калиброванные, для любого типа детектора, основанный на измерении детектор дисперсии в зависимости от интенсивности для постоянного освещения19, например, отражающей поверхности металла или сушеные краситель решение. Смещение может быть определена путем измерения скорость счета для образца без возбуждения света. Выполнив линейную регрессию детектор связанные дисперсии 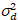 против интенсивности (я) сюжет, S и могут быть определены19:
против интенсивности (я) сюжет, S и могут быть определены19:
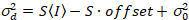 . (14)
. (14)
Наконец яркость кросс корреляции вычисляется в каждом пикселе и определяется в целом как21
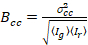 , (15).
, (15).
где 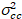 является кросс дисперсии
является кросс дисперсии 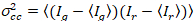 .
.
Чтобы отфильтровать долгоживущих колебания, все ccN & B расчеты выполняются после крытые вагоны, фильтрация, независимо для каждого пикселя22. Вкратце, ni, ε,я (я = g, r) и Bcc рассчитываются в скольжения сегментов например, 8-15 кадров. Таким образом полученные значения можно затем усредняются для получения последней точки номер и яркости.
Стехиометрия анализ
Для того чтобы оценить стехиометрии белковых комплексов в ячейке контактов, молекулярные яркости могут анализироваться отдельно в каждом спектральных канала для sFCCS или ccN & B данных. В sFCCS одно значение яркости получается за измерения в каждом канале. В ccN b & B гистограмма яркости всех пикселей, соответствующий контакт ячеек получается и значение средняя (или средний) может использоваться как представитель яркость для измерения. Выполняя такой же анализ на мономерных ссылку, все значения яркости могут быть нормированы непосредственно получить средний олигомерных состояние обнаруженного белковых комплексов. На данный момент важно исправить на наличие не флуоресцентные FPs, что может привести к недооценке олигомерных государства. Обычно это выполняется путем измерения яркости гомо димерной ссылкой белка23,–24 с помощью одно цвет sFCS или номер и яркость (N & B).
