Många biologiska processer inträffar på platser av cell-cell interaktioner, t.ex., cell-cell adhesion1,2,3, cell-cell fusion4 och cellulära erkännande5. Sådana händelser är särskilt viktigt under utvecklingen av flercelliga organismer och för cell-cell kommunikation, exempelvis under immunsvar. Dessa processer är typiskt medierad av proteiner som är lokaliserade på ytan, dvs på plasmamembranet (PM) på angränsande celler och genomgå särskilda interaktioner i cell cell kontakt som är tydligt reglerad i utrymme och tid. I många fall, dessa interaktioner är direkt homo- eller heterotypic protein-protein trans interaktioner, men kan också innebära joner eller ligander egenskap extracellulära linkers1. Även om det är av grundläggande betydelse, finns det brist på analyser sondera dessa specifika protein-protein interaktioner direkt i den ursprungliga miljön i levande celler. Många metoder antingen kräva cell störningar (t.ex. biokemiska analyser såsom co-immunoprecipitation6), fixering (t.ex. vissa super-upplösning optisk mikroskopi tekniker och elektronmikroskopi av cell-cell kontakter7), eller är icke-specifika, t.ex. aggregering / vidhäftning analyser8,9. För att lösa problemet, har fluorescens tekniker genomförts utifrån fluorescens resonans energi överföring (bandet)10 eller fluorescens komplettering11. Men för att uppnå tillräckligt små avstånd mellan fluorophores, kräver dessa metoder fluorescerande etiketterna på den extracellulära sidan av proteiner10, potentiellt störa trans interaktioner.
Här presenterar vi alternativa fluorescens-baserat test för protein-protein interaktioner på cell-cell kontakter. Denna strategi kombinerar fluorescens cross-korrelation strategier (skanning fluorescens cross-korrelation spektroskopi (sFCCS), cross-korrelation nummer och ljusstyrka (ccN och B)) och blandning av celler som uttrycker en fusion konstruktion av proteinet av intresse, t.ex. en vidhäftning-receptorn. De undersökta receptorerna i två samverkande cellerna är märkta med två spektralt separerade fluorescerande proteiner (FPs), från den intracellulära (se figur 1A).
De sysselsatta metoderna baseras på den statistiska analysen av fluorescens fluktuationer inducerad av diffus rörelse av fluorescerande fusionsproteinerna genom fokal volymen av en confocal laser skanning Mikroskop. Mer i detalj sonder analysens samtidig diffusionen av proteinerna av intresse för båda angränsande PMs på cell-cell kontakter. Om proteinerna genomgå trans interaktioner, kommer dessa trans komplex bära fluorescerande proteiner avger i båda spektrala kanaler, orsakar korrelerade fluorescens fluktuationer i både utsläppsländerna. Däremot, om ingen bindning uppstår, blir de antal svängningarna av proteiner i inför PMs oberoende, orsakar inga korrelerade fluktuationer. Förvärv kan utföras på två sätt: 1) sFCCS är baserat på en rad-formade scan hela cell cell kontakten och effektivt sonder samspelet i en plats som ligger i regionen kontakt. Genom en temporal analys av fluorescens svängningar ger sFCCS också dynamics information, dvs Diffusionskoefficienterna av proteinkomplex; (2) ccN & B bygger på en pixel-wise analys av en sekvens av bilder som förvärvats på cell-cell kontakt regionerna. Den har förmåga att söka och karta interaktioner längs hela kontakta region (i ett fokalplan), men ger inte information om dynamics. Båda metoderna kan kombineras med en analys av molekylär ljusstyrka, dvs genomsnittliga fluorescens signal som avges i tidsenheten av enda diffuserande proteinkomplex och, således ge uppskattningar av stökiometri av proteinkomplex på cell-cell kontakter.
I denna artikel tillhandahåller vi detaljerade protokoll för provberedning, instrumentkalibrering, datainsamling och analys att utföra presenterade analysen på en kommersiell confocal laserscanning mikroskopet. Experimenten kan utföras på något instrument utrustat med photon counting eller analoga detektorer och ett mål med hög numeriska bländaröppningen. Vi har ytterligare diskutera kritiska steg i protokollet och ger korrigering system för flera processer som orsakar tingsliga signal fluktuationer, exempelvis detektor buller, fotoblekning eller cell rörelse. Utvecklades ursprungligen för att undersöka samspelet mellan vidhäftande celler, analysen kan ändras för suspension celler eller anpassad till modellen membransystem, t.ex. giant unilamellar blåsor (GUVs) eller giant plasma membranet blåsor (GPMVs), så att den kvantifiering av interaktioner i olika lipid miljöer eller i avsaknad av en organiserad cytoskelettet12,13.
Scanning fluorescens cross-korrelation spektroskopi är en modifierad version av fluorescens cross-korrelation spektroskopi14 och utformades sond långsam diffus dynamics i lipid membran15. Den är baserad på en linje scan förvärv vinkelrätt till PM som innehåller de fluorescerande proteinerna av intresse. För att undersöka samspelet mellan två olika märkta proteinet arter, utförs förvärvet i två spektrala kanaler med två laserlinjer och två upptäckt windows för spektralt separerade fluorophores. På grund av långsam diffusion dynamiken av proteiner i PM (D≤ ~ 1 µm2per sekund), en cross-talk-gratis mätning kan utföras av omväxlande excitation systemet från linje till linje15. Analysen börjar med: 1) en justering algoritm korrigera för laterala cell rörelse utifrån block-wise genomsnitt ~ 1000 rader, 2) bestämning av placerar med maximal fluorescens signaldvs PM position, i varje block och 3) skiftande av alla block till ett gemensamt ursprung12,15, separat i varje kanal. Ett automatiskt val av pixlar motsvarar PM utförs sedan genom att välja regionen centrala från en Gaussisk passform av summan av alla anpassade linjer (dvs center ± 2.5σ). Integration av signalera i varje rad ger membran fluorescens tidsserierna F(t) i varje kanal (g = grön kanal, r = röda kanalen). Observera att pixelstorlek måste vara liten nog, t.ex. < 200 nm, att rekonstruera formen på punkten sprida funktion och hitta dess centrerar, motsvarar positionen för PM. I närvaro av betydande fotoblekning, fluorescens tidsserierna i varje kanal kan modelleras med en dubbel-exponentialfunktionen och sedan korrigeras med följande formel:16
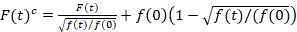 . (1)
. (1)
Det är viktigt att notera att denna formel effektivt korrigerar både amplituder och diffusion gånger erhållits från korrelation analys av F(t)c, jämfört med parametern uppskattningar som skulle erhållas från den okorrigerade F(t). Sedan funktionerna auto – och cross-korrelation (ACFs / CCFs) av fluorescensen signaler beräknas:
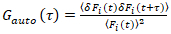 , (2).
, (2).
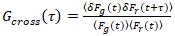 , (3).
, (3).
där δFjag = Fjag(t) – 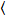 Fjag(t)
Fjag(t)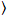 och jag = g, r.
och jag = g, r.
En tvådimensionell diffusion modell monteras sedan alla korrelation funktioner (CFs):
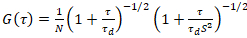 . (4)
. (4)
Här, betecknar N antalet fluorescerande proteiner i observation volym och τd diffusion tiden för varje kanal. Denna modell tar hänsyn att beskrivs i experimentell inställningen diffusion av proteiner i PM uppstår i x-z planet, i motsats till den vanliga konfigurationen av fluorescens korrelation spektroskopi (FCS) experiment på membran sondering diffusion i x-y-planet av confocal volym17. Midjan w0 och struktur faktor S, som beskriver töjning wz av fokal volymen i z, S = wz/w0, erhålls från en punkt FCS kalibrering mätning utförs med spektralt liknande färgämnen och samma optiska inställningar använder redan tillgängliga värden för den diffusionskoefficienten Dfärgämne:
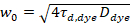 , (5).
, (5).
där τd, färgämne är den uppmätta genomsnittliga diffusion tid av färgämne molekylerna, erhållits från passande modell för tredimensionellt diffusion till data, med hänsyn till konto övergångar av en bråkdel T alla N molekyler till en triplett stat med en tidskonstant ττ:
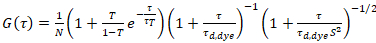 . (6)
. (6)
Slutligen beräknas Diffusionskoefficienterna (D), molekylär ljusstyrkevärden (ε) och den relativa cross-korrelationen av sFCCS data (rel.cc.) enligt följande:
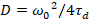 , (7).
, (7).
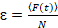 , (8).
, (8).
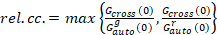 , (9).
, (9).
där Gcross(0) är amplituden av funktionen cross-korrelation och 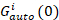 är amplituden av autokorrelation funktion i jag-th kanal.
är amplituden av autokorrelation funktion i jag-th kanal.
Denna definition av den relativa cross-korrelationen, dvs med max istället för menar i ekvation 9, tar hänsyn till att det maximala antalet komplex av två protein arter vid olika koncentrationer begränsas av den arter som finns i ett lägre nummer.
Cross-korrelation nummer och ljusstyrka är baserad på en stund analys av fluorescensintensiteten för varje pixel i en bildstapel förvärvade med tiden till en fast position i urvalet, vanligtvis bestående av ~ 100-200 ramar, med två spektrala kanaler () g = grön kanal, r = röda kanalen). Från temporal medelvärdet jagjag och variansen 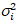 , de molekylära ljusstyrka εi och nummer njag beräknas i varje pixel och spektrala kanal (jag = g, r)18:
, de molekylära ljusstyrka εi och nummer njag beräknas i varje pixel och spektrala kanal (jag = g, r)18:
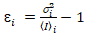 , (10).
, (10).
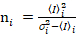 . (11)
. (11)
Det är viktigt att Observera att de givna ekvationerna gäller för det ideala fallet av en sann fotonräknande detektor. För analoga detektionssystem tillämpas följande ekvationer19,20:
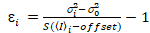 , (12).
, (12).
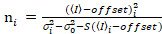 . (13)
. (13)
Här, S är konverteringsfaktorn mellan upptäckta fotoner och inspelade digitala räkningarna, 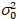 är utslaget buller och offset hänvisar till detektorn intensitet offset. Generellt, dessa kvantiteter bör kalibreras, för någon Detektortyp, baserat på mätning detektor variansen som en funktion av intensitet för jämn belysning19, t.ex. en reflekterande metallyta eller torkade dye lösning. Förskjutning kan bestämmas genom mätning räkna räntan för ett prov utan excitation ljus. Genom att utföra en linjär regression av detektor-associerade variansen
är utslaget buller och offset hänvisar till detektorn intensitet offset. Generellt, dessa kvantiteter bör kalibreras, för någon Detektortyp, baserat på mätning detektor variansen som en funktion av intensitet för jämn belysning19, t.ex. en reflekterande metallyta eller torkade dye lösning. Förskjutning kan bestämmas genom mätning räkna räntan för ett prov utan excitation ljus. Genom att utföra en linjär regression av detektor-associerade variansen 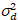 kontra intensitet (jag) tomt, S och kan vara beslutsamt19:
kontra intensitet (jag) tomt, S och kan vara beslutsamt19:
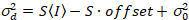 . (14)
. (14)
Slutligen, cross-korrelation ljusstyrkan beräknas i varje pixel och definieras i allmänhet som21
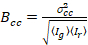 , (15).
, (15).
där 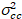 är cross-variansen
är cross-variansen 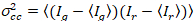 .
.
För att filtrera långlivat fluktuationer, utförs alla ccN & B beräkningar efter en boxcar filtrering, självständigt för varje pixel22. Kort, ni, εjag (jag = g, r) och Bcc beräknas i glidande segment av t.ex., 8-15 bildrutor. Erhållits på detta sätt kan vara då medelvärdet för att få den slutliga pixeln nummer och ljusstyrka värden.
Stökiometri analys
För att uppskatta stökiometri av proteinkomplex på cell-cell kontakter, kan molekylär ljusstyrkan analyseras separat för varje spektrala kanal för sFCCS eller ccN & B data. I sFCCS erhålls ett intensitetsvärde per mätning i varje kanal. I ccN & B, ett ljusstyrka histogram över alla pixlar motsvarar den cell-cell kontakten erhålls och värdet genomsnitt (eller median) kan användas som representativa ljusstyrka för mätning. Genom att utföra samma analys på en monomer referens, kan alla ljusstyrkevärden normaliseras för att direkt få den genomsnittliga Oligomera delstaten de upptäckta proteinkomplex. Vid denna punkt, är det viktigt att korrigera för förekomst av icke-fluorescerande FPs som kan leda till en underskattning av Oligomera staten. Detta görs vanligtvis genom att mäta ljusstyrka en homo-dimeriskt referens protein23,24 använder en färg sFCS eller nummer och ljusstyrka (N & B).
