The system used is an off the shelf small animal perfusion heart system but was heavily modified for use with a rabbit heart. The modifications were primarily to increase the bore size of all the tubing to assure adequate flow delivery to the rabbit heart. Great care was made to assure, at the perfusion pressures used, the flow rate of the native perfusion system exceeded the flow with heart attached by at least 5-fold. 2–12 µm pore membrane filters were placed in parallel between the fluid pump and the aortic preload bubble trap chamber to remove any debris from the heart.
Transmitted Light from the Rabbit Heart
Figure 1 presents the spectrum of the catheter (Figure 1A) and the raw spectrum of the transmitted light from the rabbit heart free wall (Figure 1B). These data reveal a very large attenuation of light in the blue region of the spectrum, however the bands of absorbance from myoglobin and the mitochondrial cytochromes can be directly observed between 490 and 580 nm in the insert. It is important in these studies to assure enough transmitted light is detected in the region from 490 to 630 nm to obtain information on the metabolically responsive cardiac chromophores. The positioning of the external and internal fibers is adjusted prior to saving data to maximize light intensity but not saturate the detector in the 625 nm region.
Reference Reduced minus Oxidized Spectra of Reference Chromophores in the heart.
Figure 2 presents the reference spectra used to fit the difference spectra collected in these studies. These references include myoglobin, cytochrome aa3 (alternatively cytochrome a605 and cytochrome a607, depending on the type of perturbation22), cytochrome a580, cytochrome bL, cytochrome bH, cytochrome c, cytochrome c1, FAD, an absorbance representation of the incident light (denoted I0, which is used to account for sieved light, that is, photons that went through the tissue without being absorbed), and a line (with varying slope and intercept to account for scattering, not shown in Figure 2). Some spectra are noisy, as the concentration of the pure reference material was very low22.
Time course of reference spectra fits during total experiment
Figure 3 represents the time course of a typical experiment calculated in step 5.15 of the protocol. This consists of a control phase, followed by a cyanide injection phase, followed by a cyanide washout, followed by a deoxygenation phase, and finally ischemia. The changes in the individual chromophores (myoglobin, cytochrome aa3, and cytochrome c) over time are plotted over time along with coronary flow rate. The optical density change of each chromophore is estimated by multiplying the fit coefficient obtained from the linear least squares routine and the representative peak of the chromophore (or the maximum absorbance of said chromophore). For example, for myoglobin, the fit coefficient of the myoglobin reference is multiplied by the value of the myoglobin reference spectrum at 580 nm. Note the rapid oxygenation of myoglobin to the addition of cyanide is matched by the increase in flow but is before the significant reduction of cytochromes. This effect is partially recovered with the washout of cyanide. Finally, full reduction of cytochromes and deoxygenation of myoglobin is obtained with ischemia. These data demonstrate that dynamic data concerning the metabolic status of the heart can easily be obtained with this methodology. The position of the spectra used for the difference spectra are marked on this time course as: C baseline, CN cyanide injection, CNW cyanide Washout, H N2 Hypoxia (nitrogen being bubbled in the perfusate instead of oxygen), and HI No flow ischemia (no perfusate flowing through the heart).
Difference Spectrum of Cyanide Treatment versus Control and Fit of Cyanide Difference Spectrum from Rabbit Heart.
To obtain a difference spectrum, two absolute spectra are subtracted. Each absolute spectrum is obtained by taking an average of many (typically 100) spectra to optimize signal to noise ratio. Figure 4A represents the difference spectrum of the control (C) and cyanide (CN) treated heart. Using the reference spectra outlined in Figure 2, the fit spectrum is calculated. The residual spectrum is the subtraction of the fit from the raw data. The same scheme is used for all the subsequent spectral presentations. Figure 4B presents the spectra amplitudes of the reference spectra (shown in Figure 2) used to fit Figure 4A. Strong increases in absorbance of most of the cytochromes are observed as the flow of electrons down the cytochrome chain was blocked by cyanide in the steady state. In addition, the absorbance of oxygenated myoglobin increased as the consumption of oxygen was eliminated by cyanide. Figure 4C presents the difference spectra and fit of the difference spectrum from CNW and CN, revealing the partial reversal of the cyanide effect. This was accomplished by selecting time points in protocol step 5.18, moving T0 to the CNW and T1 to CN region of the time course. Figure 4D presents the difference spectrum of HI and C, which represents the fully deoxygenated and reduced state of the cytosol and mitochondria versus the control condition. Again, this was performed at protocol step 5.18, moving T0 to C and T1 to HI.
Initial time course of cyanide effects on coronary flow and chromophores
Figure 5A shows an example of the initiation of the cyanide effect on the tissue. The fits for myoglobin, cytochrome a605 and cytochrome c along with the coronary flow are presented for a single heart. These time courses were created at protocol step 5.15 for the cyanide experiment. The individual difference versus the baseline (position 1) are shown in Figure 5B. The spectra were generated from the corresponding position number (1-4) on the time course. This was accomplished at protocol step 5.18, where T0 was always in position 1, and subsequently different spectra (2–4) were created by moving T1 to position 2, 3, and 4 respectively. Somewhat surprising was the observation that flow and myoglobin oxygenation increased before significant changes in cytochrome redox state. The initiation of the changes in flow and chromophore absorbance were estimated by linear extrapolation of the initial rate of change from the baseline. Using this approach, and setting the change in coronary flow as time zero, the increase in myoglobin oxygenation initiated 1.71 min ± 0.39 min after the change in flow, while cytochrome a605 and cytochrome c absorbance were nearly identical but much slower at 4.24 min ± 0.76 min and 4.34 min ± 0.77 min, respectively (n = 8). These data suggest that cyanide relaxes vascular tone24 before a major change in cardiac muscle metabolic state occurs. This effect is likely caused by cyanide encountering the vascular smooth muscle prior to reaching effective dose around the cardiac myocytes.
Estimates of Myoglobin oxygenation in control hearts
Using the cyanide data as an estimate of total myoglobin oxygenation and the ischemia data for fully deoxygenated myoglobin, we estimate that under control conditions myoglobin was only 88.2% ± 1.0% (n = 10) oxygenated, consistent with prior studies20,21,25.
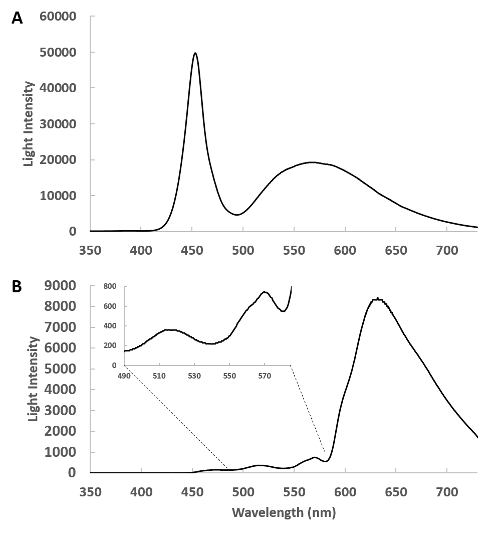
Figure 1: Spectra of the side-firing optical catheter. (A) This is a spectrum of the emitted light from the remote light source through the catheter detected with the pickup fiber at about 1 cm from the catheter. In this geometry, the heart is absent and the intensity of the light source is tuned so that the detector does not saturate. (B) The side-firing catheter is inserted in the left ventricle and the transmitted light from the heart is collected and shown. The insert shows the 400 to 580 nm region expanded, revealing the complex transmission of light from this region. Please click here to view a larger version of this figure.
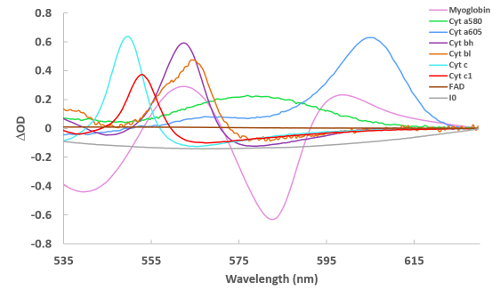
Figure 2: Reference spectra of cardiac chromophores used for spectral fitting. Spectra were collected via a variety of methods22 and are of reduced – oxidized (for the cytochromes) and deoxygenated – oxygenated (for myoglobin). For I0, the spectrum in Figure 1A is simply converted to an absorbance term to make the reference. Please click here to view a larger version of this figure.
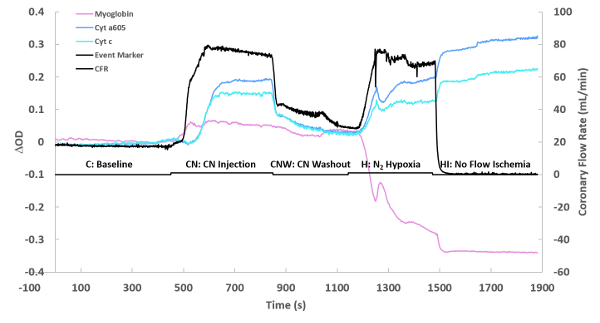
Figure 3: Flow and optical changes over time. The optical density change (ΔOD) of each chromophore is simply the fitted individual chromophore’s spectrum at its maximum absorbance. The maximum absorbance frequencies were as previously described20,26. The presented time course is for one experiment, showing a baseline, followed by cyanide injection (0.10 mM at maximum perfusate flow), cyanide washout, nitrogen hypoxia performed by replacing oxygen with nitrogen, and then complete ischemia. Please click here to view a larger version of this figure.
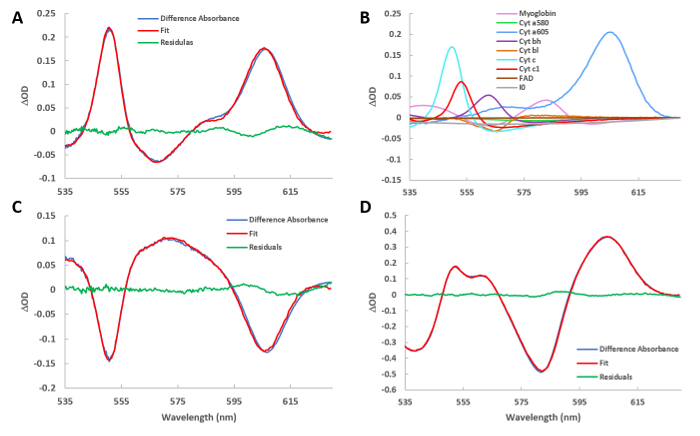
Figure 4: Fitted difference spectra of various conditions. (A) Spectrum of the cyanide injection minus the baseline. The fit spectrum obtained from the least square routine is also plotted. The residual spectrum is the difference between the raw and fit spectra. (B) The reference spectra used to create the fit presented in Figure 4A. The program scales the references in figure 2 to their relative contribution in the current difference spectrum. (C) Same as in A, but showing the difference spectrum of washout versus cyanide injection. (D) Same as in A, but showing the difference spectrum of ischemia versus baseline. Please click here to view a larger version of this figure.
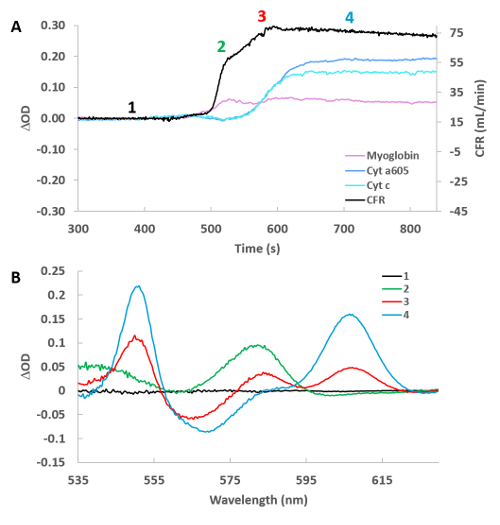
Figure 5: High temporal resolution of cyanide infusion effect on selected cytochromes, myoglobin and cardiac flow. (A) Time course of cardiac flow, deoxymyoglobin, reduced cytochrome a605, and reduced cytochrome c. Numbers refer to the position of the spectra taken relative to baseline in Figure 5B. (B) Difference spectra for the 4 positions labeled in Figure 5A versus control (position 1). Please click here to view a larger version of this figure.
