Evnen til at ingeniør genomet af alle levende organisme har mange biomedicinske og bioteknologiske applikationer, såsom korrektion af sygdom-forårsager mutationer, opførelsen af præcise cellulære modeller for sygdom undersøgelser eller generation af landbruget afgrøder med ønskværdige egenskaber. Siden århundredeskiftet, forskellige teknologier er udviklet til genom engineering i pattedyrceller, herunder meganucleases1,2,3, zink finger nukleaser4,5, eller transkription aktivere-lignende effektor nukleaser (TALENs)6,7,8,9. Men disse tidligere teknologier er enten svært at programmet eller kedelig at samle, dermed hæmmer deres udbredt vedtagelse i forskningen og industrien.
I de seneste år, de klynger interspaced regelmæssigt korte palindromiske gentagelser (CRISPR) – CRISPR-forbundet (Cas) system er opstået som en kraftfulde nye genom engineering technology10,11. Oprindeligt en adaptive immunsystem i bakterier, det har været succesfuldt implementeret for genom ændring i planter og dyr, herunder mennesker. Den primære årsag, hvorfor CRISPR-Cas har opnået så stor popularitet på så kort tid er der det element, der bringer de vigtigste Cas endonuklease, såsom Cas9 eller Cas12a (også kendt som Cpf1), til den korrekte placering i genomet er blot et kort stykke af kimære enkelt guide RN A (sgRNA), som er ligetil at design og billigt at syntetisere. Efter at blive rekrutteret til mål-webstedet, Cas enzym fungerer som et par af molekylære sakse og kløver det bundne DNA med sin RuvC, HNH eller Nuc domæner12,13,14. Den resulterende dobbelt strandede pause (DSB) er efterfølgende repareres af celler via enten ikke-homologe ende sammenføjning (NHEJ) eller homologi-instrueret reparation (HDR) pathway. I mangel af en reparation skabelon, er DSB repareret af den fejlbehæftede NHEJ vej, der kan give anledning til pseudo-tilfældige indsættelse eller deletion af nukleotider (indels) på webstedet cut, potentielt forårsager frameshift mutationer i gener, protein-kodning. Men i nærværelse af en donor-skabelon, der indeholder de ønskede DNA ændringer, DSB er repareret af high fidelity HDR pathway. Almindelige typer af donor skabeloner omfatter enkeltstrenget oligonukleotider (ssODNs) og plasmider. Førstnævnte bruges typisk, hvis de påtænkte DNA ændringer er små (f.eks. ændring af en enkelt basepar), mens sidstnævnte er typisk bruges hvis man ønsker at indsætte en relativt lang sekvens (f.eks den kodende sekvens af en grøn fluorescerende proteiner eller Normal god landbrugspraksis) i target locus.
Liv1975 aktivitet af Cas protein kræver tilstedeværelsen af et protospacer tilstødende motiv (PAM) på målet site15. PAM Cas9 er ultimo 3′ protospacer, mens PAM Cas12a (også kaldet Cpf1) er på 5′-enden i stedet16. Cas-guide RNA komplekse er ude af stand til at indføre en DSB, hvis PAM er fraværende17. Derfor, PAM placerer en begrænsning på de genomisk steder hvor en særlig Cas nukleasen er stand til at kløve. Heldigvis, Cas nukleaser fra forskellige bakteriearter typisk udviser forskellige PAM krav. Derfor, ved at integrere forskellige CRISPR-Cas systemer i vores engineering værktøjskasse, kan vi udvide antallet af websteder, der kan være målrettet i en genom. Desuden kan en naturlig Cas enzym manipuleret eller udviklet sig til at genkende alternative PAM sekvenser, yderligere udvide anvendelsesområdet for genomisk mål tilgængelige for manipulation18,19,20.
Selv om flere CRISPR-Cas systemer er tilgængelige for genom engineering formål, har de fleste brugere af teknologien påberåbt sig hovedsageligt Cas9 nukleasen fra Streptococcus pyogenes (SpCas9) af flere årsager. Først, det kræver en forholdsvis simpelthen NGG PAM i modsætning til mange andre Cas proteiner, der kan kun kløver i overværelse af mere komplekse PAMs. For det andet, det er den første Cas liv1975 implementeres med succes i humane celler21,22,23,24. For det tredje er SpCas9 langt de bedst karakteriserede enzym til dato. Hvis en forsker ønsker at bruge en anden Cas nukleasen, vil han eller hun ofte være uklart, om hvordan man bedst at designe eksperimentet og hvor godt andre enzymer vil udføre i forskellige biologiske sammenhænge i forhold til SpCas9.
For at skabe klarhed for den relative resultater af forskellige CRISPR-Cas systemer, har vi for nylig udført en systematisk sammenligning af fem Cas endonucleases-SpCas9, Cas9 enzymet fra Staphylococcus aureus (SaCas9), Cas9 enzymet fra Neisseria meningitidis (NmCas9), Cas12a enzymet fra Acidaminococcus sp. BV3L6 (AsCas12a), og Cas12a enzymet fra Lachnospiraceae bakterie ND2006 (LbCas12a)25. Vi vurderet til en fair sammenligning, de forskellige Cas nukleaser bruger det samme sæt af målwebsteder og andre forsøgsbetingelser. Undersøgelsen også afgrænset design parametre for hver CRISPR-Cas-system, som ville tjene som en nyttig reference for brugere af teknologien. Her, for bedre at kunne sætte forskerne til at gøre brug af CRISPR-Cas-system, vi leverer en trinvis protokol for optimal genom engineering med forskellige Cas9 og Cas12a enzymer (Se figur 1). Protokollen ikke kun omfatter eksperimentelle detaljer men også vigtige Designovervejelser at maksimere sandsynligheden for en vellykket genom engineering resultatet i pattedyrsceller.
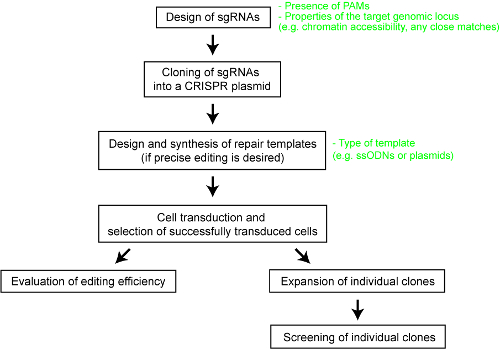
Figur 1 : En oversigt over arbejdsproces til at generere genom redigeret menneskelige cellelinjer. Venligst klik her for at se en større version af dette tal.
