We first performed the experiment to analyze phagocytosis by mouse primary AMs. Throughout all analyses, we compared AMs isolated from WT and TRIM72KO mice. As shown in Figure 1A, fluorescence microscopy revealed that phagocytosis of FITC-glass beads by mouse primary AMs occurs after 1 h of incubation. Figure 1B shows the analysis of phagocytosis by flow cytometry. The quantification of phagocytosis measured by microscopy and flow cytometry is represented in Figure 1C and Figure 1D, respectively. FcγR- and CR-mediated phagocytosis by MH-S cells is represented in Figure 2A, and Figure 2B shows the quantification. These results show that the expression of TRIM72 in MH-S cells resulted in a more than fivefold decrease in complement phagocytosis. Representative images of Alexa Fluor-488-conjugated zymosan-A particle ingestion by primary AMs isolated from WT or TRIM72KO mice are shown in Figure 2C, and the quantification is presented in Figure 2D. In vivo phagocytosis results are represented in Figure 3. Figure 3A shows differential staining identifying the presence of AMs, neutrophils, and lymphocytes, and GFP+ phagocytic cells. The percentage of BALF cells and the quantification of phagocytosis is represented in Figure 3B and Figure 3C, respectively. The percentage of body weight loss in mice after the intratracheal administration of a sublethal dose of P. aeruginosa is shown in Figure 4A, and the percentage of survival of mice at day 2 after a lethal dose of P. aeruginosa is indicated in Figure 4B. Figure 4C shows a scatter plot for the whole-lung bacterial burden at death or at day 2 after the P. aeruginosa infection in mice.
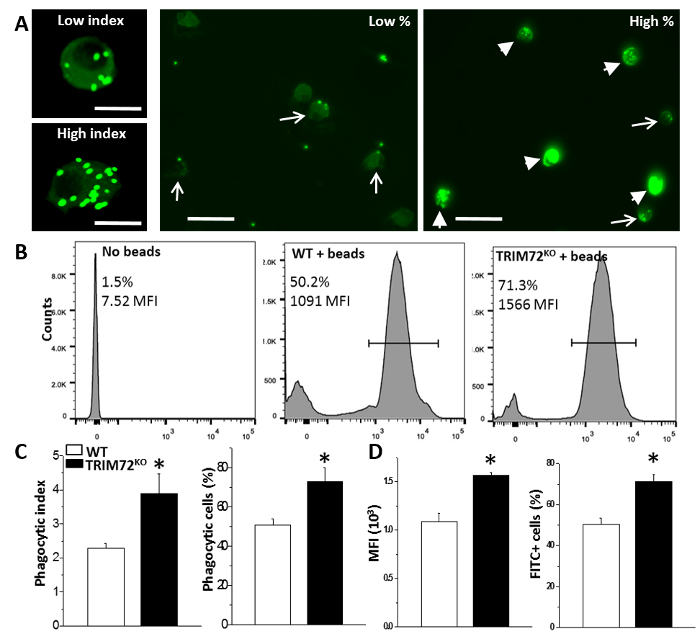
Figure 1: Phagocytosis in mouse primary alveolar macrophages. (A) Representative images of low versus high phagocytic indexes showing primary AMs containing green fluorescent beads (left); representative images showing low and high percentages of phagocytic AMs. Arrows: low phagocytic index AMs; arrowheads: high phagocytic index AMs. The scale bar = 25 µm for the left two images and 50 µm for the right two images. (B) Representative flow cytometry detection of phagocytizing cells in no-beads control, WT + beads, and TRIM72KO + beads AMs. The bars define the bead-containing cell population (in percentage) and the mean fluorescence intensity (MFI). (C) Statistics of the average phagocytic index and the percentage of phagocytic AMs in WT and TRIM72KO AMs; n = 5 for both groups, *p < 0.05. (D) Statistics of flow cytometry MFI and the percentage of FITC+ cells in WT and TRIM72KO AMs; n = 3 for both groups, *p < 0.05. This figure is reprinted with permission of the American Thoracic Society13. Please click here to view a larger version of this figure.
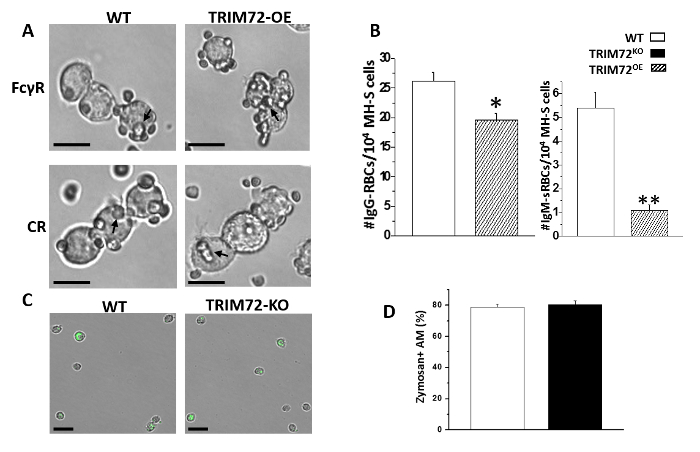
Figure 2: FcγR- and CR-mediated phagocytosis. (A) Representative images of opsonized sheep red blood cells' (SRBCs) phagocytosis by MH-S cells. The arrows point to some SRBCs. The scale bar = 25 µm. (B) Quantification of SRBCs phagocytosis by MH-S cells overexpressing TRIM72 (TRIM72OE) in the presence of IgG (FcγR-mediated phagocytosis) or IgM (CR-mediated phagocytosis); n = 6 for each group, *p < 0.05 and **p < 0.005 compared with WT control. (C) Representative images of Alexa Fluor-488-conjugated zymosan particle ingestion by primary AMs isolated from WT or TRIM72KO mice. The arrows in the panel A and B indicate zymosan + cells. The scale bar = 50 µm. (D) Statistical results of the percentage of zymosan-containing AMs; n = 3 for each group, p > 0.05. This figure is reprinted with permission of the American Thoracic Society13. Please click here to view a larger version of this figure.
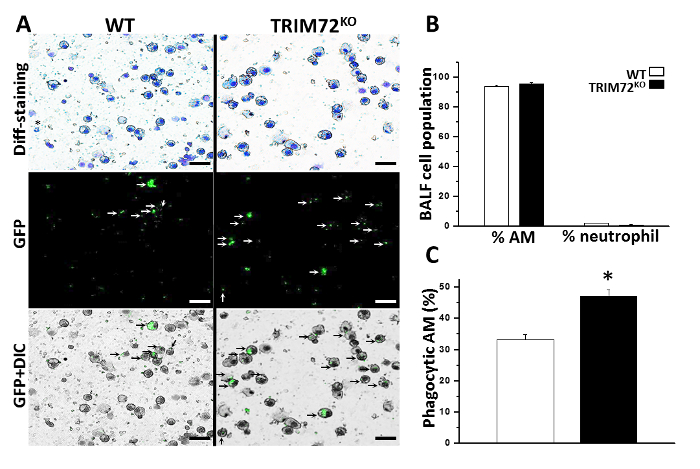
Figure 3: In vivo phagocytosis of P. aeruginosa GFP. (A) Representative images of bronchoalveolar lavage fluid (BALF) cell cytospin slides from WT and TRIM72KO mice 1 h after the injection of P. aeruginosa GFP. Kwik-Diff staining identifies AMs (large, round cells) and neutrophils; GFP identifies phagocytic cells (white arrows) and GFP+ differential interference contrast (DIC) identifies internalized GFP bacteria (black arrows) in AMs. The scale bars = 50 µm. (B) Quantification of the percentage of AMs and neutrophils in BALF of WT and TRIM72KO mice 1 h after the P. aeruginosa injection. (C) Quantification of the percentage of phagocytic AMs in WT and TRIM72KO mice; n = 3 for each group, *p < 0.05. The data are presented as mean (± SEM). This figure is reprinted with permission of the American Thoracic Society13. Please click here to view a larger version of this figure.
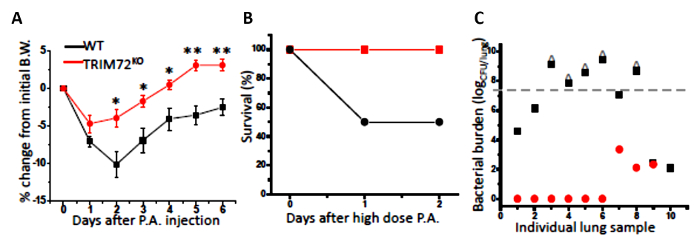
Figure 4: In vivo bacteria clearance using P. aeruginosa (P.A.). (A) The percentage of body weight (B.W.) loss of naive WT and TRIM72KO mice after the first intraperitoneal injection of 2.5 x 105 CFU/mL PAO1 (a clinical isolate of P. aeruginosa); n = 13 for WT (black squares), n = 8 for TRIM72KO (red circles), *p < 0.05, **p < 0.005 compared with WT. (B) The percentage of survival at day 2 after the 3 x 107 CFU/mL P. aeruginosa intraperitoneal injection; n = 10 for WT (solid black circles) and TRIM72KO (solid red squares), *p < 0.05 for WT versus TRIM72KO groups. (C) A scatter plot of the whole-lung bacterial burden at day 2 of the P. aeruginosa injection in WT and TRIM72KO. The gray dashed line designates the injected bacterial dose; ^ designates mice who have died. For WT versus TRIM72KO groups, p < 0.05. This figure is reprinted with permission of the American Thoracic Society13. Please click here to view a larger version of this figure.
