La transcriptase inverse amplification génique (PCR) a été l’étalon-or pour le dosage de l’ARNm. Deux essais, digital PCR (dPCR)1 et quantitative, real time PCR (qPCR)2 sont actuellement utilisés. De ces deux techniques PCR, dPCR a une plus grande sensibilité que qPCR suggérant qu’il pourrait être utilisé pour mesurer l’abondance d’ARNm dans les cellules. Toutefois, dans nos mains, dPCR analyse des ARNm de faible abondance dans les piscines de 5 à 10 ovocytes par chaque échantillon expérimental a produit des données avec faible reproductibilité et de la forte variation3. C’est probablement à cause de l’erreur expérimentale, associé à l’extraction de l’ARN et l’efficacité de la transcriptase inverse. Séquençage de RNA a également effectué à l’aide d’une seule souris et les ovocytes humains4,5. Cette technique requiert des étapes d’amplification de cDNA requis pour la génération de la bibliothèque qui augmente probablement la variabilité au sein d’un groupe expérimental. En outre, les transcriptions de faible abondance ne peuvent pas être détectables. Bien que les prix de séquençage ont baissé ces dernières années, il peut encore être coût prohibitif en raison du coût élevé des analyses bioinformatiques. Enfin, localisation de l’ARNm est un processus dynamique avec des changements spatiaux contribuant à la fonction de protéine6. Par conséquent, nous partîmes à adopter une technique qui produirait des mesures quantitatives précises et reproductibles et localisation des différents mRNAs dans les ovocytes unique.
ADN ramifié couplé à l’hybridation in situ fluorescence amplifie le signal de fluorescence plutôt qu’amplification RNA/cDNA habilitante détection seul ARNm dans les cellules individuelles 7,8,9. L’analyse est réalisée à travers une série d’hybridation, amplification (en utilisant l’ADN ramifié) et fluorescence d’étiquetage afin d’amplifier le signal de fluorescence7étapes. La technique commence par liaison de paires de sonde oligonucléotide de 18 à 25 la base qui sont complémentaires à un8,3,d’ARNm spécifiques10. Quinze à vingt paires de sonde sont conçues pour chaque spécificité de veiller à ce que de transcription pour la transcription de la cible. L’hybridation de l’ARNm spécifique est suivie d’un préamplificateur et amplificateur des sondes qui forment une configuration ramifiée. Environ 400 étiquette fluorophores lier à chaque amplificateur, résultant en une 8000-fold augmentation de fluorescence permettant la détection des différents mRNAs (Figure 1)11.
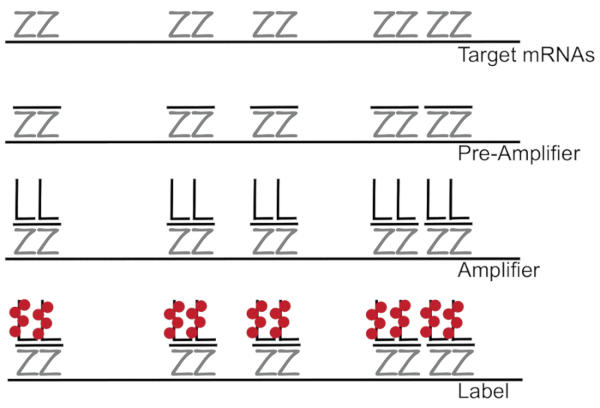
Figure 1 : schéma du protocole SM-poisson. Séquentielle hybridation d’une sonde spécifique de transcription, ramifiée amplificateur ADN et fluorophore vers une cible apparaît à l’ARNm. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
Des études antérieures utilisant des molécules simples fluorescence in situ hybridation (SM-FISH) localisée β-actine ARNm dans différents neurones12 et virus du papillome humain ADN dans le cancer du col utérin cellule lignes7. Les logiciels Spot trouver et Tracking Program identifie chaque signal fluorescent ponctuée et a été utilisé avec succès pour quantifier le nombre des ARNm dans chaque cellule3,13.
Selon les résultats de détection de l’ARNm dans les neurones12, nous avons supposé que SM-poisson s’avérera un outil utile pour quantifier les niveaux de transcription en murines ovocytes et d’embryons, y compris les ARNm de faible abondance. Cependant, la technique est optimisée pour une utilisation avec les cellules adhérentes fixes et formaldéhyde fixé paraffine intégrée des sections de tissu (FFIP). Ovocytes ne peuvent adhérer à une diapositive, même lorsqu’ils sont revêtus de la Poly-L-lysine. En outre, ils sont plus fragiles que les cellules somatiques et des sections de tissu ce qui entraîne la lyse des cellules lorsqu’elles sont soumises à certains des tampons exclusifs dans les kits disponibles dans le commerce3. Pour surmonter ces défis, les ovocytes ont été fixe et transférées manuellement entre les gouttes des tampons. En outre, tampons perméabilisation et de lavage dans les kits ont été remplacés pour réduire la lyse cellulaire. Sondes prédéfinis sont achetées aux côtés de la trousse de poissons ou des relevés de notes spécifiques peuvent être demandés. Chaque propriétaire de sonde est disponible dans l’un des trois canaux de fluorescence (C1, C2 et C3) permettant de multiplexage. Dans l’expérience actuelle, murins ovocytes étaient chiffrés à l’aide d’une sonde de C2 Nanog et une sonde de C3 Pou5f1 et double marquage. Ces sondes ont été choisis sur l’expression signalée de Nanog et Pou5f1 d’ovocytes et d’embryons. À l’issue de la procédure d’hybridation, ovocytes ont été placés en gouttes de supports de montage de l’anti-fondu pour application à histologiques. Images confocales ont été utilisées pour quantifier le nombre de signaux fluorescents ponctuées, qui représentent différents mRNAs. En plus de la quantification de l’ARNm, imagerie a également montré la distribution spatiale de l’ARNm spécifique dans la cellule, quelles autres méthodes de quantification ARN sont incapables d’atteindre. Cette technique s’est avérée pour avoir faible variabilité au sein d’un groupe expérimental permettant l’utilisation de plus petits nombres d’ovocytes dans chaque groupe expérimental afin d’identifier des différences significatives entre les groupes expérimentaux3.
