Transkriptas polymeras-kedjereaktion (PCR) har varit den gyllene standarden för mRNA kvantifieringsmetoden. Två analyser, digital PCR (dPCR)1 och kvantitativa, real time PCR (qPCR)2 används för närvarande. Av de två PCR-teknikerna har dPCR större känslighet än qPCR tyder på att det kunde användas att mäta mRNA överflöd i enstaka celler. Dock i våra händer producerat dPCR analys av låg överflöd mRNA i pooler av 5 till 10 ägg per varje experimental prov data med låg reproducerbarhet och hög variation3. Detta beror sannolikt på den experimentella fel i samband med RNA-extraktion och omvänd transkription effektivitet. RNA-sekvensering har också utförts med ett enda musklick och mänskliga äggceller4,5. Denna teknik kräver cDNA förstärkning steg krävs för bibliotek produktion vilket sannolikt ökar variationen inom en experimentell grupp. Låg överflöd avskrifter kan dessutom inte upptäckas. Även om sekvensering priserna har gått ner de senaste åren, kan det fortfarande vara kostnadseffektivt oöverkomliga på grund av de höga kostnaderna för bioinformatik analyser. Slutligen, mRNA lokalisering är en dynamisk process med rumsliga förändringar bidrar till protein funktion6. Därför har vi ställt ut för att anta en teknik som skulle ge exakta och reproducerbara kvantitativa åtgärder och lokalisering av enskilda mRNA i enda oocyter.
Grenad DNA kopplat till fluorescens in situ hybridisering förstärker fluorescens signal i stället för ljudförstärkande RNA/cDNA möjliggör detektering av enda mRNA i enskilda celler 7,8,9. Analysen utförs genom en serie av hybridisering, amplifiering (med förgrenade DNA) och fluorescens märkning steg för att förstärka fluorescens signal7. Tekniken börjar med bindningen av 18 – till 25-base oligonukleotiden sonden par som kompletterar en specifik mRNA3,8,10. Femton till tjugo sonden par är utformade för varje avskrift säkerställa specificitet för målet avskriften. MRNA-specifika hybridisering följs av förförstärkare och slutsteg sonder som bildar en förgrenad konfiguration. Ungefärligt binder 400 etikett fluorophores till varje förstärkare, vilket resulterar i en 8000-fold ökning fluorescens som möjliggör identifiering av enskilda mRNA (figur 1)11.
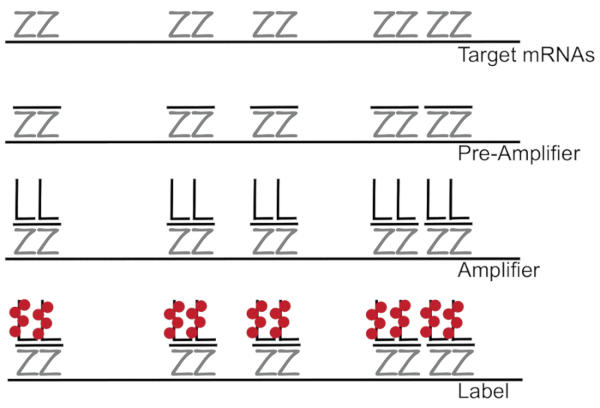
Figur 1: Schematisk av protokollet SM-fisk. Sekventiell hybridisering av avskrift specifika sond, grenad DNA förstärkare och fluorophore till ett mål som mRNA visas. Klicka här för att se en större version av denna siffra.
Tidigare studier med enda molekyl fluorescens i situ hybridisering (SM-fisk) lokaliserade β-aktin mRNA i enskilda nervceller12 och humant papillomvirus DNA i livmoderhalscancer cell linjer7. Det datorprogram som plats att hitta och spåra programmet identifierar enskilda punktuell fluorescerande signalen och har framgångsrikt använts för att kvantifiera antalet mRNA i varje cell3,13.
Baserat på resultaten av mRNA detection i nervceller12, hypotesen vi att SM-fisk skulle visa sig vara ett användbart verktyg att kvantifiera avskrift nivåer i murina oocyter och embryon inklusive låg överflöd mRNA. Men tekniken är optimerad för användning med vidhäftande fast celler och formaldehyd fast paraffin inbäddade (FFPE) vävnadssnitt. Oocyter kan inte följa en bild, även när de är belagda med Poly-L-Lysin. Dessutom är de mer ömtålig än somatiska celler och vävnadssnitt resulterar i cellen lysis när de utsätts för vissa av de egenutvecklade buffertarna i kommersiellt tillgängliga kit3. För att övervinna dessa utmaningar, var oocyter fast och manuellt överförs mellan droppar av buffertar. Dessutom byttes permeabilisering och tvätta buffertar i kits för att minska cellys. Fördesignade sonder köps tillsammans med fisk kit eller specifika avskrifter kan begäras. Varje egenutvecklade probe set finns i en av tre fluorescens kanaler (C1, C2 och C3) för att låta för multiplexering. I det nuvarande experimentet var murina oocyter dual-färgade och kvantifieras med hjälp av en C2 Nanog sond och en C3 Pou5f1 sond. Dessa sonder valdes baserat på rapporterade uttryck för Nanog och Pou5f1 i oocyter och embryon. Vid avslutningen av hybridisering steg placerades oocyter i droppar anti fade montering media för ansökan till histologiska bilder. Confocal bilder har använts för att kvantifiera antalet punktuell fluorescerande signaler som företräder enskilda mRNA. Förutom att kvantifiera mRNA, imaging visade också den rumsliga fördelningen av specifika mRNA i cellen, är som andra RNA kvantifiering metoder oförmögen att uppnå. Denna teknik visade sig ha låg variabilitet inom en experimentell grupp som medger användning av mindre antal ägg i varje experimentell grupp att identifiera betydande skillnader mellan experimentella grupper3.
