Некоторые активные виды кислорода (ROS) способны окислять двойные углеродные связи баз ДНК и некоторые углероды в дезоксирибозе, создавая окисленные базы и разрывы ДНК-нити1. В качестве отрицательно заряженной молекулы, богатой атомами азота и кислорода, ДНК также является мишенью для электрофильных групп, которые ковалентно реагируют с нуклеофильных участков (азота и кислорода), давая продукты, которые называются ДНК аддуктов2. Таким образом, ДНК-аддукты и окисленные базы ДНК являются примерами повреждений ДНК, которые являются полезными биомаркерами для оценки токсичности веществ, которые электрофильные, генерировать реактивные электрофилы при биотрансформации, или индуцировать окислительный стресс1, 2. Хотя модифицированные базы ДНК могут быть удалены из ДНК с помощью базы или эксцизии нуклеотидов (BER или NER), индукция дисбаланса между генерацией и удалением поражений ДНК в пользу первого приводит к чистому увеличению их уровней в сверхурочных ДНК3 . Результаты являются увеличение скорости мутации ДНК, снижение экспрессии генов, и уменьшилась активность белка2,4,5,6,7, эффекты, которые тесно связаны с развития болезней. Мутации ДНК могут влиять на различные клеточные функции, такие как клеточная сигнализация, клеточный цикл, целостность генома, стабильность теломер, эпигеном, Структура хроматина, РНК-сплайсинг, протеиновый гомеостаз, метаболизм, апоптоз и клеточная дифференциация8 ,9. Стратегии, направленные на замедление темпов мутации клеток и развитие хронических заболеваний (например, рак, нейродегенеративные заболевания) проходят через знание источников мутации, среди них, поражения ДНК и их причины.
ROS генерируемые эндогенно в избытке, из-за воздействия загрязнителей, стойкие воспаления, патофизиологии болезни (например, диабет) и т.д., являются важными причинами био повреждения, включая ДНК и повреждение липидов1. В качестве примера, Высокореактивные гидроксильные радикальные (OH) формируется из H2O2 уменьшение переходных ионов металлов (Fe2 +, Cu+) окисляет ДНК баз, ДНК сахара Моисея и полиненасыщенных жирных кислот при диффузии контролируемых ставки10. Среди 80 уже охарактеризовано окисленные нуклеобазы3, наиболее изученное из них-8-оксо-7, 8-дигидрогуанин (8-оксигуа) или 8-оксо-7, 8-diгидро-2′-дезоксигуаньозин (8-Окродго, рис. 1 клетки млекопитающих10,11. Он формируется моноэлектронным окислением гуанина, или гидроксильных радикалами или сингзным кислородной атакой гуанина в ДНК1. Полиненасыщенные жирные кислоты являются другими важными целями высокореактивных окислители, такие как •Oh, которые инициируют процесс перекисного окисления липидов1,12. Он порождает гидропероксиды жирных кислот, которые могут разлагаться на электрофильные альдегиды и эпоксидииды, такие, как малондиалдегид, 4-гидрокси-2-нонеал, 2, 4-decadienal, 4, 5-эпоксидной-(2е)-decenal, гексенал, акролейн, кротональдегид, которые являются способны образовывать мутагенное экзоцикловую ДНК-аддукты, такие как малондиалдегид-, пропано-, или этено аддукс1,12,13. Этено-аддукты 1,n2-этоно-2′-дезокгугуаньозин (1,n2-в., рис. 1) и 1,n6-Этоно-2′-дезоксиаденозин (1,n6-, на рисунке 1 ) были предложены в качестве потенциальных биомаркеров в патофизиологии воспаления14,15.
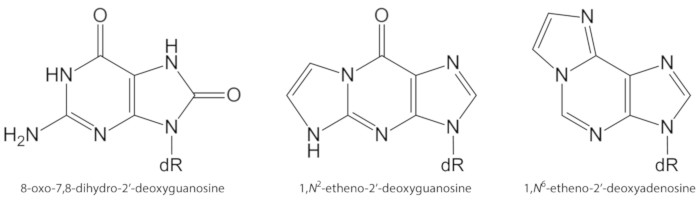
На рисунке 1. В настоящем исследовании количественно определены химические структуры повреждений ДНК. dR = 2 ́-дезоксирибоза. Эта цифра была изменена с Оливейра et al.34. Пожалуйста, нажмите здесь, чтобы посмотреть увеличенном варианте этой фигуры.
Исследования, проведенные в начале 1980-х годов, позволили чувствительное обнаружение 8-Окродго высокой производительностью жидкой хроматографии, связанной с электрохимическим обнаружением (HPLC-РДРВ). Количественное определение 8-окродго путем HPLC-РДРВ в нескольких биологических системах, подвергшихся окислительной конъюнкции, привело к распознаванию 8-окродго как биомаркера оксидативно индуцированного базового повреждения в ДНК1,16. Несмотря на прочность и возможность количественной оценки 8-oxodGuo в низком диапазоне17, HPLC-РДРВ измерений полагаться на точность анализатор времени удержания для анализа идентификации и на разрешение хроматографии, чтобы избежать помех других составляющих выборки. Поскольку для электрохимического обнаружения требуется использование соли (например, фосфата калия, ацетат натрия) на подвижной фазе, поддержание адекватных аналитических условий требует регулярной очистки времени и оборудования.
С другой стороны, использование бактериального ДНК репарации фермента формаламидина ДНК гликозилазы (ФПГ) и, впоследствии, человеческий 8-оксигуанин гликозиназы 1 (hOGG1), для обнаружения и удаления 8-Oxoгуа из ДНК, возникла как способ для индукции ДНК щелочных лабильных Сайтов. Щелочные лабильные узлы преобразуются в разрывы в ДНК и позволяют очень высоко чувствительным косвенным количественному измерению 8-oxoGua путем щелочного электрофореза одноклеточного геля (“анализ кометы”). Высокая чувствительность и выполнение анализов без необходимости клеточной экстракции ДНК являются основными преимуществами этого типа анализа. Он дает самые низкие устойчивые-государственные уровни 8-oxoGua в ДНК, как правило, 7-10 раз ниже, чем уровни, полученные биоаналитическими методами, основанными на HPLC. Однако, это косвенное измерение 8-oxogua и некоторые недостатки являются отсутствие специфики или неизвестной эффективности ремонтных ферментов, используемых1,16,18.
Иммуноанализы представляют собой другой набор методов, используемых для обнаружения 8-Оксикогуа1 и экзоцикликс ДНК-аддуктов, таких как 1,n6-Дадо и 1,n2-ДГО12. Несмотря на чувствительность, недостатком использования антител для выявления повреждений ДНК является отсутствие специфики из-за перекрестной реактивности других компонентов биологических образцов, в том числе нормальных баз ДНК1,12. ДНК экзоциклика, в том числе 1,n6-Дадо и 1,n2-ДГО, также могут быть обнаружены и количественно по высокочувствительным 32P-postlabeling анализ12. Высокая чувствительность 32P-постмаркировки позволяет использовать очень небольшое количество ДНК (например, 10 мкг) для обнаружения около 1 аддода на 1010 нормальных баз19. Однако использование радиохимикатов, отсутствие химической специфичности и низкая точность являются некоторыми недостатками19,20.
Общим ограничением указанных выше методов является низкая избирательность или специфичность для обнаружения желаемых молекул. В этом сценарии, HPLC в сочетании с электрораспылителем ионизации тандем масс-спектрометрии (HPLC-ЭСИ-МС/МС и HPLC-MS3) превратилась в золотой стандарт для количественной оценки модифицированных нуклеолы в биологических матрицах, таких как ДНК, моча, плазма и слюна 1 , в 19 , 20. преимуществами HPLC-ЭСИ-МС/МС являются чувствительность (как правило, в низком диапазоне фмоль) и высокая специфичность, предоставляемая i) Хроматографическое разделение, II) Характеристика и известный образец фрагментации молекул внутри массы спектрометр столкновение камеры, и III) точное измерение выбранной массы для зарядки соотношение (m/z) в нескольких режиме мониторинга реакции1,19. Польза изотопно чистых маркированные внутренне стандарты добавляет преимущество коррекций для потерь молекулы во время этапов гидролиза дна и анализата обогащения, также, как для разницы анализирована ионизации между образцами. Он также помогает в определении правильного хроматографического пика, когда более чем один пик присутствует1,12,19,20.
Несколько методов, основанных на хплс-Нези-МС/МС были использованы для количественной оценки 8-окродго, 1,N6-Дадо и 1,N2-ДГО в ДНК, извлеченные из различных биологических образцов12,15,20 ,21,22,23,24,25,26,27,28,29 . Мелкие частицы (PM2,5) несут органические и неорганические химические вещества, такие как полициклические ароматические углеводороды (ПАУ), нитро-ПАУ, альдегиды, кетоны, карбокфильные кислоты, хинолинов, металлы и водорастворимые ионы, которые могут вызывать воспаление и окислительный стресс, условия, благоприятствуют возникновению био повреждений и заболеваний30,31,32,33. Мы представляем здесь подтверждены HPLC-НЕЗИ-МС/МС методы, которые были успешно применены для количественной оценки 8-oxodGuo, 1,N6-Дадо и 1,N2-ДГО в легких, печени и почек ДНК A/J мышей для Оценка воздействия окружающего PM2,5 экспозиции34.
