Algunas especies reactivas de oxígeno (ROS) son capaces de oxidar enlaces dobles de carbono de bases de ADN y algunos carbonos en la fracción desoxirribosa, generando bases oxidadas y roturas de hebras de ADN1. Como una molécula cargada negativamente rica en átomos de nitrógeno y oxígeno, el ADN es también un objetivo para los grupos electrófilos que reaccionan covalentemente con los sitios nucleófilos (nitrógeno y oxígeno), dando productos que se denominan aductos de ADN2. Por lo tanto, los aductos de ADN y las bases de ADN oxidado son ejemplos de lesiones de ADN que son biomarcadores útiles para la evaluación de la toxicidad de sustancias que son electrofilicas, generan electrófilos reactivos sobre la biotransformación, o inducen el estrés oxidativo1, 2. Aunque las bases de ADN modificadas se pueden eliminar del ADN por la reparación de la escisión de base o nucleótido (BER o NER), la inducción de un desequilibrio entre la generación y la eliminación de las lesiones de ADN en favor de la primera conduce a un aumento neto de sus niveles en el ADN horas extras3 . Los resultados son el aumento de las tasas de mutación del ADN, la reducción de la expresión génica y la disminución de la actividad proteica2,4,5,6,7, efectos estrechamente relacionados con el desarrollo de enfermedades. Las mutaciones de ADN pueden afectar diversas funciones celulares, como la señalización celular, el ciclo celular, la integridad del genoma, la estabilidad del telómero, el epigenoma, la estructura de la cromatina, el empalme de ARN, la homeostasis proteica, el metabolismo, la apoptosis y la diferenciación celular8 ,9. Las estrategias para ralentizar las tasas de mutación celular y el desarrollo de enfermedades crónicas (p. ej., cáncer, enfermedades neurodegenerativas) pasan por el conocimiento de las fuentes de mutación, entre ellas, las lesiones de ADN y sus causas.
Los ROS generados endógenamente en exceso, debido a la exposición a los contaminantes, la inflamación persistente, la fisiopatología de la enfermedad (p. ej., diabetes), etc., son causas importantes del daño de la biomolécula, incluyendo el ADN y el daño lipídico1. Por ejemplo, el radical hidroxilo altamente reactivo (OH) formado a partir de la reducción de H2O2 por iones metálicos de transición (fe2 +, cu+) oxida las bases de ADN, la fracción de azúcar de ADN y los ácidos grasos poliinsaturados en la difusión controlada tarifas10. Entre las 80 nucleobasas oxidadas ya caracterizadas3, el más estudiado es 8-oxo-7, 8-dihydroguanine (8-oxoGua) o 8-oxo-7,8-dihydro-2′-desoxyguanosine (8-oxodGuo, figura 1), una lesión que es capaz de inducir transversiones gt en células de mamíferos10,11. Se forma por la oxidación electrónica mono de guanina, o por el ataque de oxígeno de la guanina a radical hidroxilo o singlete en el ADN1. Los ácidos grasos poliinsaturados son otros objetivos importantes de los oxidantes altamente reactivos, tales como •Oh, que inician el proceso de peroxidación lipídica1,12. Da lugar a hidroperóxidos de ácidos grasos que pueden descomponerse a aldehídos electrófilos y epoxialdehídos, tales como malondialdehído, 4-hidroxi-2-Nonenal, 2,4-Decadienal, 4, 5-epoxi-(2E)-decenal, hexenal, Acrolein, Crotonaldehído, que son puede formar aductos de ADN exocíclico mutagénico, tales como malondialdehído-, propano-, o los aductos de etheno1,12,13. Los aductos de etheno 1,n2-etheno-2′-deoxyguanosine (1,n2-εdguo , figura 1) y 1,n6-etheno-2′-deoxyadenosine (1,n6-εdado, figura 1 ) se han sugerido como biomarcadores potenciales en la fisiopatología de la inflamación14,15.
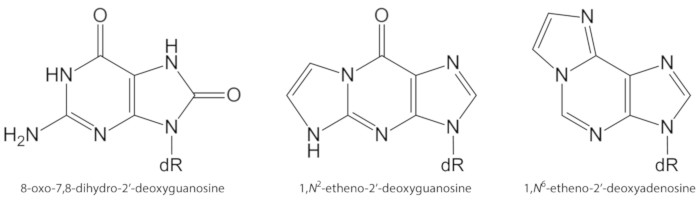
Figura 1. Estructuras químicas de las lesiones de ADN cuantificadas en el presente estudio. dR = 2 ́-desoxiribosa. Esta cifra ha sido modificada de Oliveira et al.34. Por favor, haga clic aquí para ver una versión más grande de esta figura.
Los estudios realizados a comienzos de la década de 1980 permitieron la detección sensible de 8-oxodGuo por cromatografía líquida de alto rendimiento acoplada a la detección electroquímica (HPLC-ECD). La cuantificación de 8-oxodguo por HPLC-ECD en varios sistemas biológicos sometidos a condiciones oxidantes condujo al reconocimiento de 8-oxodguo como un biomarcador de daño base inducido oxidativamente en el ADN1,16. Aunque son robustos y permiten la cuantificación de 8-oxodGuo en la gama baja de fmol17, las mediciones de HPLC-ECD se basan en la precisión del tiempo de retención del analito para la identificación del analito y en la resolución de la cromatografía para evitar interferencias de otros componentes de la muestra. Como la detección electroquímica requiere el uso de sal (por ejemplo, fosfato de potasio, acetato de sodio) en la fase móvil, el mantenimiento de condiciones analíticas adecuadas necesita tiempo de limpieza de columna y equipo de rutina.
Alternativamente, el uso de la enzima de reparación de ADN bacteriológico formamidopirimidina ADN glicosilasa (FPG) y, después, 8-oxoguanina glucosilasa 1 (hOGG1), para la detección y eliminación de 8-oxogua de ADN, surgió como una manera de la inducción de ADN alcalino lábil Sitios. Los sitios del lábil alcalino se convierten en roturas de la hebra de ADN y permiten la cuantificación indirecta muy alta sensible de 8-oxogua por electroforesis de gel de una sola célula alcalina (“ensayo de cometa”). La alta sensibilidad y la realización de los análisis sin necesidad de extracción de ADN celular son las principales ventajas de este tipo de ensayo. Da los niveles de estado estacionario más bajos de 8-oxoGua en el ADN, típicamente 7-10 veces más bajo que los niveles obtenidos por métodos bioanalíticos basados en HPLC. Sin embargo, es una medida indirecta de 8-oxogua y algunos inconvenientes son la falta de especificidad o la eficiencia desconocida de las enzimas de reparación utilizadas1,16,18.
Los inmunoensayos son otro conjunto de métodos utilizados para la detección de aductos de ADN 8-oxoGua1 y exocíclico, tales como 1,n6-dado y 1,n2-dguo12. A pesar de la sensibilidad, una deficiencia del uso de anticuerpos para la detección de lesiones de ADN es la falta de especificidad debido a la reactividad cruzada a otros componentes de muestras biológicas, incluyendo las bases de ADN normales1,12. Los aductos de ADN exocíclico, incluyendo 1,n6-DAdo y 1,n2-dguo, también pueden ser detectados y cuantificados por los ensayos altamente sensibles de 32P-postetiquetado12. La alta sensibilidad de 32P-el etiquetado posterior permite el uso de cantidades muy pequeñas de ADN (por ejemplo, 10 μg) para la detección de aproximadamente 1 aducto por 1010 bases normales19. Sin embargo, el uso de radioquímicos, la falta de especificidad química y la baja precisión son algunas desventajas19,20.
Una limitación compartida de los métodos citados anteriormente es la baja selectividad o especificidad para la detección de las moléculas deseadas. En este escenario, la HPLC acoplada a la espectrometría de masas tándem de ionización electrospray (HPLC-ESI-MS/MS y HPLC-MS3) evolucionó como el estándar de oro para la cuantificación de nucleósidos modificados en matrices biológicas, como el ADN, la orina, el plasma y la saliva 1 , 19 , 20. las ventajas de los métodos HPLC-ESI-MS/MS son la sensibilidad (típicamente en la gama baja de fmol) y la alta especificidad proporcionada por i) la separación cromatográfica, II) el patrón característico y conocido de fragmentación molecular dentro de la masa cámara de colisión espectrómetro, y III) la medición precisa de la relación masa a carga seleccionada (m/z) en el modo de monitorización de reacción múltiple1,19. El uso de estándares internos etiquetados isotópicamente añade la ventaja de las correcciones para las pérdidas de moléculas durante la hidrólisis del ADN y los pasos de enriquecimiento del analito, así como para las diferencias de la ionización del analito entre las muestras. También ayuda en la identificación del pico cromatográfico correcto cuando hay más de un pico presente1,12,19,20.
Se han utilizado varios métodos basados en HPLC-ESI-MS/MS para la cuantificación de 8-oxodguo, 1, N6-dado y 1,n2-dguo en el ADN extraído de diferentes muestras biológicas12,15,20 ,21,22,23,24,25,26,27,28,29 . Las partículas finas (PM2,5) transportan productos químicos orgánicos e inorgánicos, tales como hidrocarburos aromáticos policíclicos (PAHs), Nitro-PAHS, aldehídos, cetonas, ácidos Carboxilicos, quinolinas, metales y iones solubles en agua, que pueden inducir inflamación y estrés oxidativo, condiciones que favorecen la ocurrencia de daño biomolecular y enfermedad30,31,32,33. Aquí presentamos métodos validados de HPLC-ESI-MS/MS que se aplicaron con éxito para la cuantificación de 8-oxodGuo, 1,N6-DAdo y 1,N2-dguo en pulmón, hígado y ADN renal de ratones A/J para la evaluación de la efectos de la exposición ambiental PM2,5 34.
