Vi beskriver en total intern reflektion fluorescens-antal och ljusstyrka (TIRF-N & B) avbildning metod för att bestämma den genomsnittliga oligomeriska tillstånd av receptor molekyler vid plasmamembranet i levande celler, som syftar till att länka receptorn församlingen dynamiken till proteiners biologiska funktion (figur 1).
Vid extracellulära ligand bindning, receptorer initiera intracellulära signaltransduktion beroende på deras konformation, oligomerisering, potentiella Co-receptorer och membran sammansättning. Trots betydelsen och ubiquity av receptor oligomerization, erkänd som en viktig händelse i cellulära signalering1,2,3,4,5,6, 7, få metoder kan upptäcka klusterhändelser och mäta graden av kluster experimentellt8,9. Den konfokalvolym (x, y ≈ 300 nm, z ≈ 900 nm) är otillräckligt löst för att bevisa molekylär interaktion och stökiometri, även efter optimering av bildrestaurering algoritmer10. Den sub-Unit sammansättningen av protein oligomerer kan inte lösas på en rent rumslig grund även av super-upplösning metoder på x, y upplösning på 20-70 nm som PALM11, Storm12, och sted13. Dessutom kan deras temporala upplösning (i storleksordningen minuter per bild) inte följa kinetik i intervallet sekunder. Single molekyl Step-blekning löser stoichiometri av protein oligomerer endast om de är orörbara14.
En av de mest mångsidiga metoderna för att mäta densitet och oligomerisering av fluorescerande märkta proteiner inom enstaka bilder är den rumsliga intensiteten fördelning analys (SpIDA), som förlitar sig på rumsliga provtagning. Den är tillämplig på både kemiskt fasta och levande celler, och gör det möjligt att analysera flera regioner av intresse i cellen samtidigt med hjälp av standardfluorescens mikroskopi15. Alternativt moment metoder, såsom fluorescens-korrelationsspektroskopi (FCS)16, Photon Counting histogram (PCH)17, och antal och ljusstyrka (N & B)18,19, är lämpliga för kvantitativa Oligomer Mätningar. Dessa metoder analyserar variationer i fluorescensintensiteten som kan observeras i tid när fluoroforerna sprids in och ut ur en belysnings volym. Amplituderna i intensiteten kan beskrivas unikt genom den molekylära ljusstyrkan hos fluorophore (ε) och det genomsnittliga antalet fluoroforer (n) inom belysnings volymen17 (figur 2). Normalt kan diffusions koefficienten för fluoroforer och det genomsnittliga antalet molekyler (omvänt besläktade med G (0)-värdet) inom belysnings volymen erhållas genom FCS20. Men eftersom diffusions tiden bara skalas med kubisk roten av massan, är FCS inte tillräckligt känslig för att upptäcka förändringar i molekylmassa21. I praktiken kan enfärgade FCS inte detektera Dimerization av membran receptorer. PCH löser blandningar av olika oligomerer noggrant. Med mer än två moment av amplitudfördelningen upptäcker den molekyler med olika ljusstyrka som upptar samma belysnings volym. Scanning FCS22 och utveckling, såsom intressant par-korrelation av molekylär ljusstyrka (pcomb) Approach23, infördes för att utvidga räckvidden för tillämpligheten av fluorescenskorrelations metoder i biologiska system24 , förbli Single Point-metoder som saknar förmåga att snabba mätningar i ett stort område av en cell, som kräver många på varandra följande observationer vid varje pixel och datainsamling i storleksordningen sekunder.
N & B är en förenklad version av pch som endast beaktar den första och andra ögonblicken av amplituden för fluorescensfördelningen, nämligen Medelintensiteten, och varians, σ2 (figur 2)18,19 och på grund av detta, kan den inte bestämma molar fraktion av okända oligomerer i en blandning, men bara uppskattar den genomsnittliga oligomerisering tillstånd av blandningen. N & B har dock fördelen av att arbeta med relativt mindre tidsserier av bilder av levande celler än PCH på en pixel-för-pixel-basis, helt enkelt genom att övervaka fluktuationerna i tiden för fluorescensintensiteten. Eftersom N & B minskar tiden per pixel till några mikrosekunder, kan det följa snabbt oligomerisering kinetik över stora cellområden, vilket gör att bild förvärv på en tidsskala av sekunder i raster skanning mikroskopi (t. ex., konfokal, 2-Photon) och millisekunder i kamerabasbaserad mikroskopi (t. ex. TIRFM).
Flera rapporter har visat förmåga N & B att kvantifiera antalet subenheter i protein kluster genom att avbilda utökade cell regioner. Paxillin-EGFP-kluster upptäcktes vid vidhäftnings platserna i CHO-K1-cellerna25, och den intracellulära aggregeringen av patogenen Httex1p peptid BESKREVS i cos-7-celler26. N & B applicerades för att följa den ligand-drivna oligomeriseringen av ErbB-receptorn27, och effekten av LIGAND FGF21 på Klothob (KLB) och FGFR1c i hela cells28. Kombinationen av tirf Imaging och N & B-analys användes för att visa att dynamin-2 i första hand är tetrameriska genom hela cellmembranet29. Vi tillämpade N & B till både raster skanning och tirf bilder för att bevisa ligand-driven Dimerization av uPAR och FGFR1 cellmembran receptorer30,31.
Fluorescenskorrelations metoder, såsom N & B, FCS och PCH, bygger på föreställningen att ockupationen av partiklar i en öppen volym följer en Poissonfördelning. Eftersom endast fotoner som avger fluoroforer kan upptäckas, är medelvärdet för en uppmätt fluorescensintensitet kontra tid i en bildpunkt i bilden  , produkten av det genomsnittliga antalet fluorofomedel i belysnings volymen, n, och deras Molekylär ljusstyrka, ε17:
, produkten av det genomsnittliga antalet fluorofomedel i belysnings volymen, n, och deras Molekylär ljusstyrka, ε17:

där ε uttrycks som det antal fotoner som avges per tidsenhet (konventionellt per sekund) per molekyl när molekylen står i centrum för belysnings volymen.
Ljusstyrka är en egenskap hos varje fluorophore i en given förvärvs uppsättning, medan intensiteten är summan av alla bidrag från alla fluoroforer. I biologiska tävlingar kommer ljusstyrkan att öka med ökningen av antalet fluoroforer som fluktuerar tillsammans, vilket ger information om oligomeriseringstillståndet hos det fluorescerande-märkta proteinet. Fluktuationsamplituderna vid en given pixel mäts från variansen för fluorescenssignalen, σ2:

Om medelvärdet av kvadraten på intensitet,  och kvadraten på medelvärdet av intensiteten,
och kvadraten på medelvärdet av intensiteten,  beräknas utifrån de individuella intensitetsvärdena i varje pixel i varje ram:
beräknas utifrån de individuella intensitetsvärdena i varje pixel i varje ram:
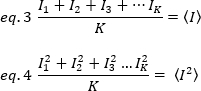
där K är antalet totala bildrutor i tidsserierna. Experimentellt är det nödvändigt att beräkna för hela bildserien variansen som beskriver scattern för de enskilda intensitetsvärdena vid varje pixel i en enskild bild runt medelintensitetsvärdet. Avvikelsen inkluderar alla variationer av olika ursprung. I en första närhet, variancen som de diffusa partiklarna i belysnings volymen, σ20, kan AVS kil ras från variancen tack vare som avkännaren sköt stojar, σ2d. De två varianserna är oberoende; således ges total avvikelsen av summan:

Variationen, på grund av molekylära variationer in och ut ur detekterings volymen, är linjärt beroende av den molekylära ljusstyrkan och intensiteten:

Omdisponering av EQ. 6 enligt EQ. 1:

Enligt det typiska konceptet i fluorescenskorrelationspektroskopi, anger ekvation 7 att variationen på grund av antalet variationer beror på kvadraten på partikel ljusstyrkan.
Därefter är variansen på grund av detektor svängningar en linjär funktion av den upptäckta intensiteten, under antagandet att detektorn drivs under dess mättnadsgräns19:

I fallet av fotonen som räknar avkännare a= 1 och c= 0, således är detektor avvikelsen jämbördig till den genomsnittliga styrkan:

För att tillämpa dessa begrepp på verkliga mätningar i Live-celler, Gratton och kollegor18 definiera skenbar ljusstyrka, B, för varje pixel som förhållandet mellan variansen över den genomsnittliga intensiteten:

B är den parameter som mäts experimentellt. I detta arbete, tidsserie bilder av FGFR1-receptorer vid plasmamembranet av HeLa celler fångas upp av TIRF mikroskopi och den genomsnittliga skenbara ljusstyrkan, B, bestäms av N & B-analys. Sedan, efter tillsats av FGF2, på varandra följande tidsserier fångas för att följa förändringarna i själv-församlingen av receptorn molekyler i membranet ytan efter stimulering av receptorn med den kanoniska ligand.
Men eftersom detektorn i TIRF-mikroskopet är en EMCCD-kamera måste uttrycket för den skenbara ljusstyrkan ändras till19:

där offset är intensitetsförskjutningen av detekterings elektroniken som är kännetecknande för detektor inställningarna. Varians och medelintensitet för en analog detektor ges respektive av:
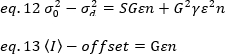
där G är den analoga förstärkningen i digitala nivåer (DL/fotoner), S, de digitala nivåerna per foton19, ges av lutningen på en intensitet kontra var Ians tomt för en ljuskälla med konstant intensitet (inga temporala fluktuationer). Γ-faktorn är relaterad till pixel detekterings volymens form. Enligt Hassler et al.32, den γ faktorn är lika med 0,3 för tirf Imaging arbetar vid maximal vinst på detekterings kameran19. Offset-, S-och G-parametrarna är kamerans och mikroskopens egenskaper. Den skenbara ljusstyrkan, B, erhålls genom omdisponering av EQ. 11 enligt EQ. 12 och 13:

Experimentellt är ε en komplex funktion av laserintensitet och detekterings effektiviteten i systemet. Eftersom B/S är linjärt beroende av ε är det dock endast viktigt att fastställa det relativa värdet av ε för ett givet detektions läge:

där ε står i proportion till ε. En kalibrering utförs fortfarande med en intern referens.
