Der er et betydeligt antal hormonforstyrrende stoffer (EDC), der er blandt de farligste stoffer i vores miljø. Disse er hovedsageligt østrogene forbindelser, der forurener vand fra naturressourcer. Den kemiske mangfoldighed af de stoffer, der tilhører gruppen, gør det vanskeligt at teste for deres tilstedeværelse, da der er behov for forskellige analysemetoder til påvisning ning af dem. Baseret på deres kemiske struktur er det meget vanskeligt at afgøre, om et stof rent faktisk er i stand til at fungere som en østrogen. Desuden er disse stoffer aldrig til stede i en ren form i miljøet, så deres virkninger kan blive påvirket af andre forbindelser, for1. Dette problem kan løses ved effekt-afsløre metoder, såsom brug af biomonitor / bioindikatororganismer,der viser østrogene effekter2,3,4,5.
For nylig, en række celle linje6 og gær-baserede testsystemer2,,3 er blevet udviklet til at opdage østrogene effekter. Men, disse er generelt kun i stand til at opdage bindingen af stoffet til østrogen receptor2,3. Desuden er de ude af stand til at modellere komplekse fysiologiske processer i organismen, eller til at opdage hormon-følsomme faser af livsstadiet; De fører således ofte til falske resultater.
Det er kendt, at visse gener reagerer følsomt på østrogen i levende organismer7. Påvisning af genprodukter ved molekylærbiologi metoder er også muligt på protein eller mRNAniveau 8,9, men normalt indebærer dyr offer. Dyreværnslovgivningen er blevet strengere, og der er en stigende efterspørgsel efter alternative testsystemer, der minimerer antallet og lidelserne hos dyr, der anvendes til forsøg eller udskiftning af dyremodellen med et andet modelsystem10. Med opdagelsen af fluorescerende proteiner og oprettelsen af biomarkørlinjer udgør transgene teknologier et godt alternativ11. Med disse linjer, aktivering af en østrogen-følsomme gen kan testes in vivo.
Blandt hvirveldyr er fiskenes potentiale i miljørisikovurderingen enestående. De tilbyder mange fordele i forhold til pattedyr modeller: at være akvatiske organismer, de er i stand til at absorbere forurenende stoffer gennem hele deres krop, producere et stort antal afkom, og nogle af deres arter er kendetegnet ved kort generation tid. Deres endokrine system og fysiologiske processer viser store ligheder med andre hvirveldyr og endda med pattedyr, herunder mennesker12.
Flere gener til påvisning af østrogene effekter i fisk er også kendt. De vigtigste er østrogen receptorer aromatase-b, choriogenin-H, og vitellogenin (vtg)7,13. For nylig, flere østrogen-producerende biosensor linjer er også blevet skabt af fisk modeller, der anvendes i laboratoriet, såsom fra zebrafisk (Danio rerio)4,5,14,15,16,17. Den største fordel ved zebrafisk i at skabe biosensor linjer er den gennemsigtige krop af embryoner og larver, fordi fluorescerende reporter signal kan derefter let studeres in vivo uden at ofre dyret10. Ud over dyrebeskyttelse er det også et værdifuldt træk, da det giver mulighed for at studere den samme persons reaktion på forskellige behandlingstider18.
Disse forsøg bruger en vitellogenin reporter transgene zebrafisk linje15. Den transgene konstruktion, der anvendes til udvikling af Tg(vtg1:mCherry) har en lang (3,4 kbp) naturlig vitellogenin-1 promotor. Østrogen receptor (ER) er en forstærker protein aktiveres af ligander, der er en repræsentant for steroid/nukleare receptor superfamily. ER binder sig til specifikke DNA-sekvenser kaldet østrogen respons elementer (EREs) med høj affinitet og transaktiverer genekspression som reaktion på estradiol og andre østrogene stoffer, så jo mere ERE i promotor forårsager enstærkere respons 19. Der er 17 ERE-lokaliteter i frem initiativtagerregionen for transgenetkonstruktionen Tg(vtg1:mCherry), og de forventes at efterligne udtrykket af det oprindelige vtg-gen15. Der er et kontinuerligt udtryk for det fluorescerende signal hos seksuelt modnede hunner. Hos hanner og embryo er udtrykket i leveren dog kun synligt ved behandling med østrogene stoffer (Figur 1).
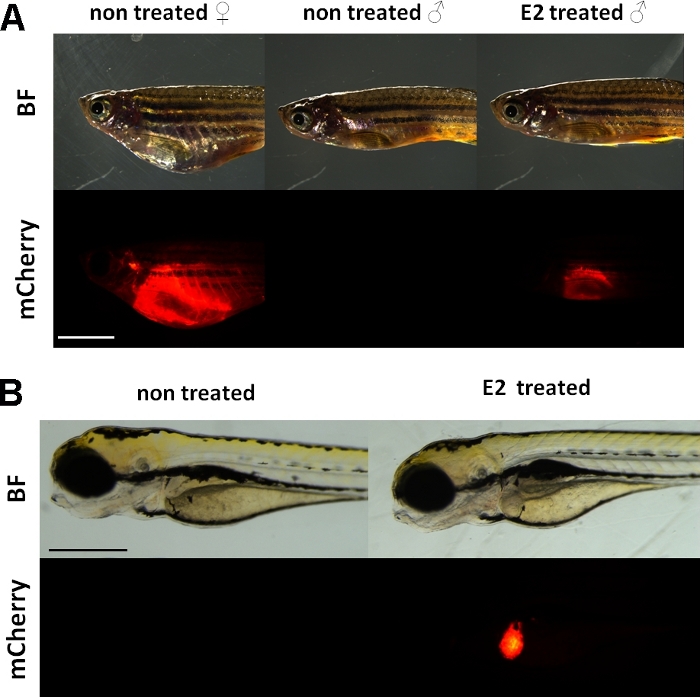
Figur 1: Rødt fluorescerende signal i leveren af vtg1:mCherry transgen voksen zebrafisk og 5 dpf embryoner efter 17-ß-estradiol (E2) induktion. Hos hun- og han- og han-patienter behandlet med E2 (25 μg/l eksponeringstid:48 timer) er der tydelig kraftig fluorescens i leveren, selv gennem den pigmenterede hud. Intet fluorescerende signal er synligt hos ubehandlede hanner (A). Efter E2 induktion (50 μg/L eksponeringstid: 0-120 hkf), kan der også observeres et rødt fluorescerende signal i leveren på 5 dpf-embryoner, som ikke er synligt i kontrolembryoner (B). Mens fluorescerende signal er kontinuerligt til stede i voksne kvinder, primært mænd og embryoner af linjen er egnet til påvisning af østrogene effekter. (BF: lyse felt, mCherry: rød fluorescerende filter visning, enkelt plain billeder, Scale bar A: 5mm, skala bar B: 250 μm) Klik her for at se en større version af dette tal.
Svarende til den endogene vitellogenin, mCherry reporter er kun udtrykt i leveren. Fordi vitellogenin kun produceres i nærvær af østrogen, der er ingen fluorescerende signal i kontrollerne. Fordi udtrykket er kun i leveren, evalueringen af resultaterne er meget lettere15.
Følsomheden og anvendeligheden af denne linjes embryoner er blevet undersøgt på forskellige østrogene sammensatte blandinger og også påmiljøprøver 15,20, og i de fleste tilfælde dosis-respons relationer blev dokumenteret (Figur 2). I tilfælde af meget giftige, hovedsagelig hepatotoksiske stoffer (f.eks. zearalenon), kan der dog kun ses et meget svagt fluorescerende signal i leveren af behandlede embryoner, og det maksimale fluorescerende signal kan nås inden for et meget lille koncentrationsområde, hvilket gør det vanskeligt at etablere dosiseffektforhold20.
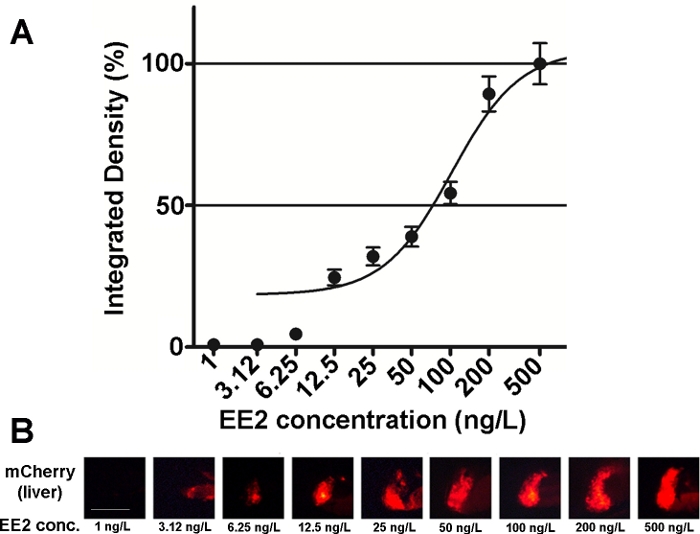
Figur 2: Dosis-respons diagram (A) og fluorescerende billeder (mCherry) af leveren (B) eksponeret for 17-α-ethynilestradiol (EE2), i 5 dpf vtg1:mCherry larver. Resultaterne udtrykkes som integreret tæthed, der genereres ud fra signalstyrken og størrelsen af det berørte område (±SEM, n = 60). 100% refererer til det observerede maksimum. Fluorescerende signalintensitet steg gradvist med koncentration. Skalabar = 250 μm. Klik her for at se en større version af dette tal.
Der er flere østrogene stoffer til stede i miljøet, 17-β-estradiol (miljøkoncentration: 0,1-5,1 ng/L)21,17-α-ethynylestradiol (miljøkoncentration: 0,16-0,2 μg/L)22, zearalenon (miljøkoncentration: 0,095–0,22 μg/L)23, bisphenol-A (miljøkoncentration: 0,45–17,2 mg/l)24. Ved testning af disse stoffer i ren aktiv form ved hjælp af transgene mCherry-embryoner var de laveste koncentrationer af observeret effekt (LOEC) til fluorescerende tegndetektion 100 ng/L for 17-ß-estradiol. 1 ng/L for 17-α-ethynilestradiol, 100 ng/L for zearalenon og 1 mg/L for bisphenol-A (96-120 hkf behandling), som er meget tæt på eller inden for området af miljømæssige koncentrationeraf stofferne 15. Den Tg (vtg1:mCherry) transgene linje kan hjælpe med at opdage østrogene i spildevand prøver efter direkte eksponering. Linjen er lige så følsom som den almindeligt anvendte gær østrogen test, den bioluminiserende gær østrogen (BLYES) analyse15. Ved hjælp af denne linje er de beskyttende virkninger af beta-cyclodextriner mod zearalenon-induceret toksicitet blevet bekræftet ved hjælp af kemiskeblandinger 20.
I en nylig rapport blev in vivo-brugen af den transgene linje påvist ved hjælp af to østrogene zearalenon (ZEA) metabolitter, α- og β-zearalenol (α-ZOL og β-ZOL)25. Protokollen baseline er hensigtsmæssigt at studere de østrogene virkninger af flere forbindelser eller miljøprøver på Tg (vtg1:mCherry) embryoner.
