Il y a un nombre important de composés perturbateurs endocriniens (EDC) qui sont parmi les substances les plus dangereuses dans notre environnement. Ce sont principalement des composés œstrogéniques qui contaminent l’eau des ressources naturelles. La diversité chimique des substances appartenant au groupe rend les tests de leur présence difficiles, car différentes méthodes d’analyse sont nécessaires pour leur détection. Sur la base de leur structure chimique, il est très difficile de déterminer si une substance est réellement capable d’agir comme un œstrogène. En outre, ces substances ne sont jamais présentes sous une forme pure dans l’environnement, de sorte que leurs effets peuvent être affectés par d’autres composés, trop1. Ce problème peut être résolu par des méthodes de détection d’effet, telles que l’utilisation d’organismes biomonitor/bioindicateurs qui présentent des effets œstrogéniques2,3,4,5.
Récemment, une variété de cellules ligne6 et de systèmes d’essai à base de levure2,3 ont été développés pour détecter les effets œstrogéniques. Cependant, ceux-ci ne sont généralement en mesure de détecter la liaison de la substance au récepteur d’oestrogène2,3. En outre, ils sont incapables de modéliser des processus physiologiques complexes dans l’organisme, ou de détecter les phases sensibles aux hormones des stades de la vie; ainsi, ils conduisent souvent à de faux résultats.
On sait que certains gènes réagissent sensiblement à l’œstrogène dans les organismesvivants 7. La détection des produits génétiques par des méthodes de biologie moléculaire est également possible au niveau de protéine ou d’ARNm8,9, mais implique généralement le sacrifice d’animaux. Les lois sur la protection des animaux sont devenues plus strictes, et il y a une demande croissante de systèmes d’essai alternatifs qui minimisent le nombre et la souffrance des animaux utilisés dans les expériences ou le remplacement du modèle animal par un autre système modèle10. Avec la découverte de protéines fluorescentes et la création de lignées de biomarqueurs, les technologies transgéniques offrent une bonne alternative11. Avec ces lignes, l’activation d’un gène sensible à l’œstrogène peut être testée in vivo.
Chez les vertébrés, le potentiel des poissons dans l’évaluation des risques environnementaux est exceptionnel. Ils offrent de nombreux avantages par rapport aux modèles de mammifères : étant des organismes aquatiques, ils sont capables d’absorber les polluants dans tout leur corps, produisent un grand nombre de descendants, et certaines de leurs espèces sont caractérisées par un temps de production de courte durée. Leur système endocrinien et leurs processus physiologiques présentent de grandes similitudes avec d’autres vertébrés et même avec les mammifères, y compris les humains12.
Plusieurs gènes pour la détection des effets œstrogéniques chez les poissons sont également connus. Les plus importants sont les récepteurs d’oestrogène aromatase-b, choriogenine-H, et vitellogénine (vtg)7,13. Récemment, plusieurs lignées de biocapteurs produisant des œstrogènes ont également été créées à partir de modèles de poissons utilisés en laboratoire, comme celui du poisson zèbre (Danio rerio)4,5,14,15,16,17. Le principal avantage du poisson zèbre dans la création de lignes de biocapteur est le corps transparent des embryons et des larves, parce que le signal de journaliste fluorescent peut alors être facilement étudié in vivo sans sacrifier l’animal10. En plus de la protection des animaux, il est également une caractéristique précieuse car il permet d’étudier la réaction d’un même individu à différents moments du traitement18.
Ces expériences utilisent un journaliste de vitellogenine transgénique zèbre ligne15. La construction transgène utilisée pour le développement de Tg(vtg1:mCherry) a un long (3,4 kbp) naturel vitellogenin-1 promoteur. Le récepteur d’oestrogène (ER) est une protéine d’exhausteur activée par des ligands qui est un représentant de la superfamille stéroïde/récepteur nucléaire. ER se lie à des séquences d’ADN spécifiques appelées éléments de réponse d’oestrogène (EER) avec une forte affinité et transactive l’expression des gènes en réponse à l’estradiol et d’autres substances œstrogéniques, de sorte que plus ERE dans le promoteur provoque une réponse plus forte19. Il y a 17 sites ERE dans la région de promoteur de la construction transgène Tg(vtg1:mCherry) et on s’attend à ce qu’ils imitent l’expression du gène vtg indigène15. Il y a une expression continue du signal fluorescent chez les femelles sexuellement matures. Cependant, chez les mâles et les embryons, l’expression dans le foie n’est visible que lors du traitement avec des substances œstrogéniques (figure 1).
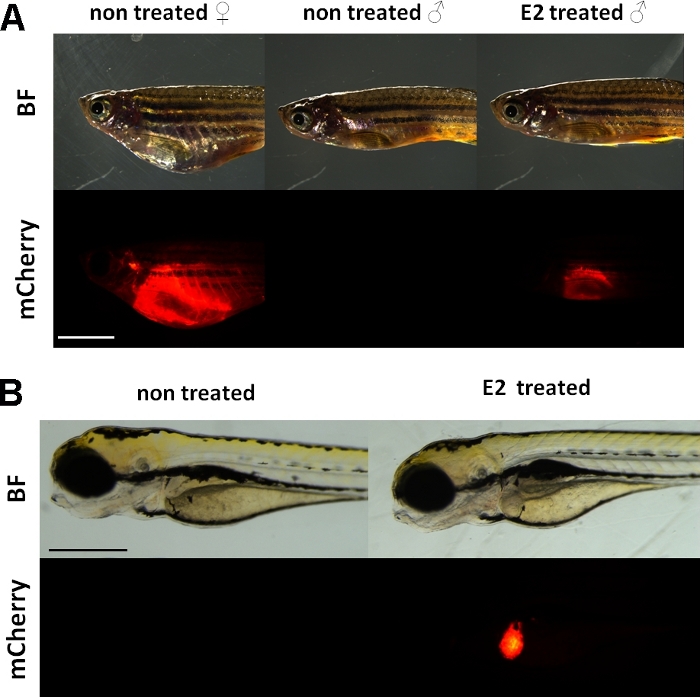
Figure 1 : Signal fluorescent rouge dans le foie du poisson zèbre adulte transgénique vtg1:mCherry et de 5 embryons de dpf, après induction de 17-ß-estradiol (E2). Chez les femelles et chez les hommes traités par E2 (25 μg/L temps d’exposition : 48hrs), une forte fluorescence du foie est visible même à travers la peau pigmentée. Aucun signal fluorescent n’est visible chez les mâles non traités (A). Après induction e2 (50 μg/L temps d’exposition : 0-120 hpf), un signal fluorescent rouge dans le foie de 5 embryons de dpf peut également être observé, ce qui n’est pas visible dans les embryons témoins (B). Bien que le signal fluorescent soit continuellement présent chez les femelles adultes, principalement les mâles et les embryons de la lignée sont appropriés pour détecter les effets œstrogéniques. (BF: champ lumineux, mCherry: vue de filtre fluorescent rouge, images simples, barre d’échelle A: 5mm, barre d’échelle B: 250 μm) Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
Semblable à la vitellogénine endogène, le journaliste mCherry n’est exprimé que dans le foie. Parce que la vitellogénine n’est produite qu’en présence d’œstrogènes, il n’y a pas de signal fluorescent dans les commandes. Parce que l’expression est seulement dans le foie, l’évaluation des résultats est beaucoup plus facile15.
La sensibilité et la facilité d’utilisation des embryons de cette lignée ont été étudiées sur divers mélanges de composés œstrogéniques ainsi que sur des échantillons environnementaux15,20, et dans la plupart des cas, des relations dose-réponse ont été documentées ( figure2). Toutefois, dans le cas de substances hautement toxiques, principalement hépatotoxiques (p. ex., zéaralenone), seul un signal fluorescent très faible peut être visible dans le foie des embryons traités et l’intensité maximale causée par le signal fluorescent peut être atteinte dans une très petite plage de concentration, ce qui rend difficile l’établissement de relations dose-effet20.
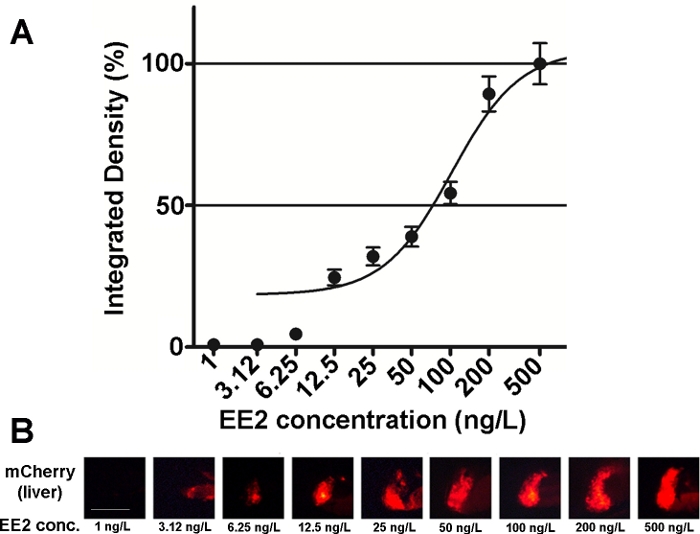
Figure 2 : Diagramme dose-réponse (A) et images fluorescentes (mCherry) du foie (B) exposés au 17-α-ethynilestradiol (EE2), dans 5 larves de dpf vtg1:mCherry. Les résultats sont exprimés sous forme de densité intégrée générée par la force du signal et la taille de la zone touchée (±SEM, n = 60). 100% se réfère au maximum observé. L’intensité du signal fluorescent a augmenté progressivement avec la concentration. Barre d’échelle = 250 μm. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
Il existe plusieurs substances œstrogéniques dans l’environnement, comme 17-β-estradiol (concentration environnementale : 0,1–5,1 ng/L)21, 17-α-éthylestradiol (concentration environnementale : 0,16–0,2 μg/L)22, zéaralenone (concentration environnementale : 0,095–0,22 μg/L)23, bisphénol-A (concentration environnementale : 0,45–17,2 mg/L)24. Lors de l’essai de ces substances sous une forme purement active à l’aide d’embryons transgéniques mCherry, les concentrations d’effet les plus faibles observées (LOEC) pour la détection des signes fluorescents étaient de 100 ng/L pour 17-ß-estradiol, 1 ng/L pour 17-α-ethynilestradiol, 100 ng/L pour la zéaralenone, et 1 mg/L pour le bisphénol-A (traitement 96–120 hpf), qui est très proche ou dans la gamme des concentrations environnementales des substances15. La ligne transgénique Tg(vtg1:mCherry) peut aider à détecter l’œstoïscité dans les échantillons d’eaux usées après une exposition directe. La ligne est aussi sensible que le test d’oestrogène de levure couramment utilisé, l’oestrogène de levure bioluminscente (BLYES) essai15. À l’aide de cette ligne, les effets protecteurs des bêta-cyclodextrines contre la toxicité induite par la zéaralénone ont été confirmés à l’aide de mélanges chimiques20.
Dans un rapport récent, l’utilisation in vivo de la lignée transgénique a été démontrée à l’aide de deux métabolites œstrogéniques de zéaralenone (ZEA), α- et β-zearalénol (α-ZOL et β-ZOL)25. La base de référence du protocole est appropriée pour étudier les effets œstrogéniques de plusieurs composés ou échantillons environnementaux sur les embryons de Tg(vtg1:mCherry).
