Det er et betydelig antall endokrine forstyrrende forbindelser (EDC) som er blant de farligste stoffene i vårt miljø. Dette er hovedsakelig estrogenic forbindelser som forurenser vann fra naturressurser. Det kjemiske mangfoldet av stoffene som tilhører gruppen gjør testing for deres tilstedeværelse vanskelig, da forskjellige analytiske metoder er nødvendig for deres deteksjon. Basert på deres kjemiske struktur er det svært vanskelig å avgjøre om et stoff faktisk er i stand til å fungere som et østrogen. I tillegg er disse stoffene aldri tilstede i ren form i miljøet, så deres effekter kan påvirkes av andre forbindelser, for1. Dette problemet kan løses ved effektdetekterende metoder, for eksempel bruk av biomonitor / bioindicator organismer som viser estrogenic effekter2,3,4,5.
Nylig, en rekke celle linje6 og gjær-baserte testsystemer2,3 har blitt utviklet for å oppdage estrogenic effekter. Disse er imidlertid vanligvis bare i stand til å oppdage bindingen av stoffet til østrogenreseptoren2,3. I tillegg er de ikke i stand til å modellere komplekse fysiologiske prosesser i organismen, eller for å oppdage hormonfølsomme faser av livsstadier; dermed fører de ofte til falske resultater.
Det er kjent at visse gener reagerer følsomt på østrogen i levende organismer7. Påvisning av genprodukter ved molekylærbiologi metoder er også mulig på protein eller mRNA nivå8,9, men vanligvis innebærer dyr offer. Dyrevernslover har blitt strengere, og det er en økende etterspørsel etter alternative testsystemer som minimerer antall og lidelser av dyr som brukes i eksperimenter eller utskifting av dyremodellen med et annet modellsystem10. Med oppdagelsen av fluorescerende proteiner og etableringen av biomarkørlinjer, gir transgene teknologier et godt alternativ11. Med disse linjene kan aktiveringen av et østrogenfølsomt gen testes in vivo.
Blant virveldyr er potensialet for fisk i miljørisikovurdering enestående. De tilbyr mange fordeler fremfor pattedyrmodeller: å være vannlevende organismer, de er i stand til å absorbere forurensende stoffer gjennom hele kroppen, produsere et stort antall avkom, og noen av deres arter er preget av kort generasjonstid. Deres endokrine system og fysiologiske prosesser viser store likheter med andre virveldyr og selv med pattedyr, inkludert mennesker12.
Flere gener for påvisning av estrogenic effekter i fisk er også kjent. Det viktigste er østrogen reseptorer aromatase-b, choriogenin-H, og vitellogenin (vtg)7,13. Nylig har flere østrogenproduserende biosensorlinjer også blitt opprettet fra fiskemodeller som brukes i laboratoriet, for eksempel fra sebrafisk (Danio rerio)4,5,14,15,16,17. Den største fordelen med sebrafisk i å skape biosensorlinjer er den gjennomsiktige kroppen til embryoer og larver, fordi det fluorescerende reportersignalet deretter lett kan studeres in vivo uten å ofre dyret10. I tillegg til dyrevern er det også en verdifull funksjon som det gjør det mulig å studere reaksjonen til samme person på forskjellige tider av behandlingen18.
Disse eksperimentene bruker en vitellogenin reporter transgen sebrafisk linje15. Transgene konstruksjonen som brukes til utvikling av Tg(vtg1:mCherry) har en lang (3,4 kbp) naturlig vitellogenin-1 arrangør. Østrogen reseptor (ER) er en enhancer protein aktivert av ligander som er en representant for steroid/kjernefysisk reseptor superfamilie. ER binder seg til spesifikke DNA-sekvenser kalt østrogenresponselementer (ERE) med høy affinitet og transaktiverer genuttrykk som svar på østradiol og andre estrogenic stoffer, slik at jo mer ERE i arrangøren forårsaker en sterkere respons19. Det er 17 ERE nettsteder i arrangørregionen av Tg (vtg1:mCherry) transgene konstruere og de forventes å etterligne uttrykket av den innfødte vtg genet15. Det er et kontinuerlig uttrykk for det fluorescerende signalet hos seksuelt modnede kvinner. Men hos menn og embryo er uttrykket i leveren bare synlig ved behandling med estrogenic stoffer (figur 1).
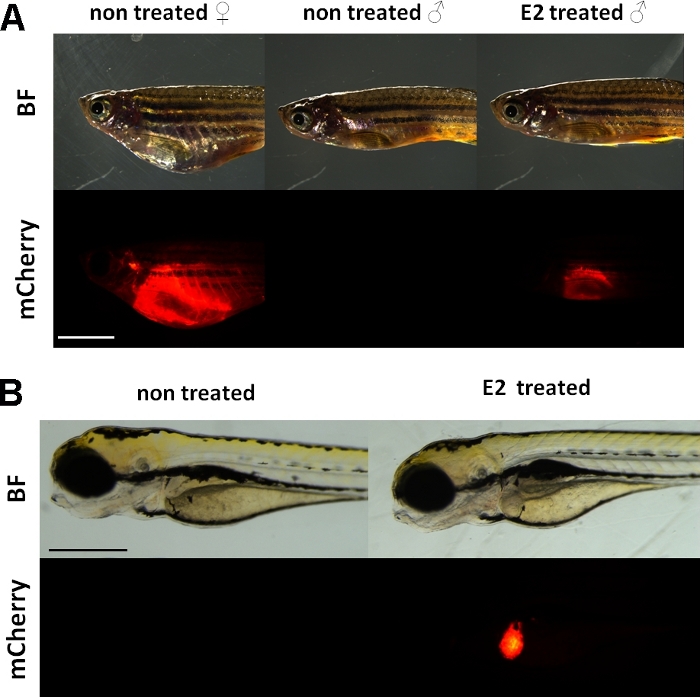
Figur 1: Rødt fluorescerende signal i leveren av vtg1:mCherry transgene voksne sebrafisk og 5 dpf embryoer, etter 17-ß-østradiol (E2) induksjon. Hos kvinner og hos hann behandlet med E2 (25 μg/l eksponeringstid:48 timer) er sterk fluorescens i leveren synlig selv gjennom den pigmenterte huden. Ingen fluorescerende signal er synlig hos ubehandlet mann (A). Etter E2 induksjon (50 μg/l eksponeringstid: 0-120 hkf), kan et rødt fluorescerende signal i leveren på 5 dpf embryoer også observeres, noe som ikke er synlig i kontrollembryoer (B). Mens fluorescerende signalet er kontinuerlig til stede hos voksne kvinner, er primært menn og embryoer i linjen egnet for å oppdage estrogenic effekter. (BF: bright field, mCherry: rød fluorescerende filtervisning, enkle slettebilder, Skala bar A: 5mm, skala bar B: 250 μm) Vennligst klikk her for å se en større versjon av dette tallet.
I likhet med den endogene vitellogenin, er mCherry reporter bare uttrykt i leveren. Fordi vitellogenin bare produseres i nærvær av østrogen, er det ingen fluorescerende signal i kontrollene. Fordi uttrykket er bare i leveren, er evalueringen av resultatene mye enklere15.
Følsomheten og brukervennligheten til denne linjens embryoer har blitt undersøkt på ulike estrogenic sammensatte blandinger og også på miljøprøver15,,20, og i de fleste tilfeller dose-respons relasjoner ble dokumentert (Figur 2). Men i tilfelle av svært giftig, hovedsakelig hepatotoksisk, stoffer (f.eks zearalenone), bare et svært svakt fluorescerende signal kan være synlig i leveren av behandlede embryoer og maksimal intensitet fluorescerende signal forårsaket kan nås innenfor et svært lite konsentrasjonsområde, noe som gjør det vanskelig å etablere dose-effekt relasjoner20.
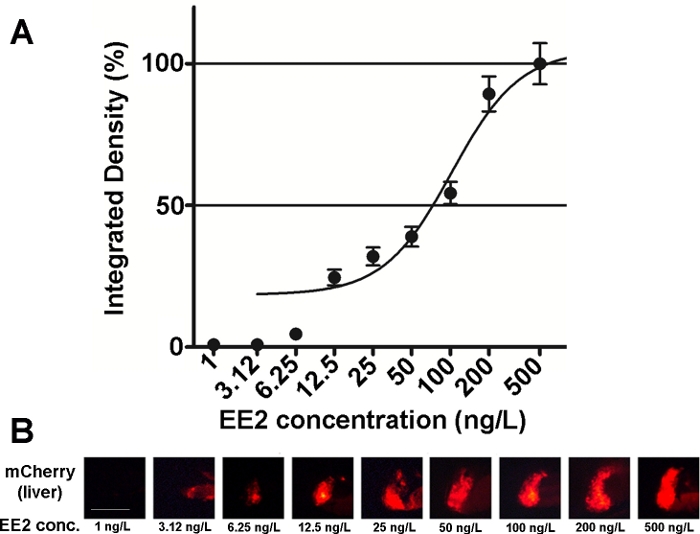
Figur 2: Doseresponsdiagram (A) og fluorescerende bilder (mCherry) i leveren (B) eksponert for 17-α-ethynilestradiol (EE2), i 5 dpf vtg1:mCherry larver. Resultatene uttrykkes som integrert tetthet generert fra signalstyrken og størrelsen på det berørte området (±SEM, n = 60). 100 % refererer til det observerte maksimumet. Fluorescerende signalintensitet økte gradvis med konsentrasjon. Skala bar = 250 μm. Klikk her for å se en større versjon av dette tallet.
Det er flere estrogenic stoffer tilstede i miljøet, som 17-β-østradiol (miljøkonsentrasjon: 0,1–5,1 ng/l)21,17-α-ethynylestradiol (miljøkonsentrasjon: 0,16–0,2 μg/L)22, zearalenone (miljøkonsentrasjon: 0.095–0.22 μg/L)23, bisfenol-A (miljøkonsentrasjon: 0.45–17.2 mg/L)24. Ved testing av disse stoffene i ren aktiv form ved hjelp av mCherry transgene embryoer var de laveste observerte effektkonsentrasjonene (LOEC) for fluorescerende tegndeteksjon 100 ng/l for 17-ß-østradiol, 1 ng/l for 17-α-ethynilestradiol, 100 ng/l for zearalenon, og 1 mg/l for bisfenol-A (96–120 hpf behandling), som er svært nær eller innenfor rekkevidden av miljøkonsentrasjoner av stoffene15. Tg(vtg1:mCherry) transgene linjen kan bidra til å oppdage estrogenicity i avløpsvannprøver etter direkte eksponering. Linjen er like følsom som den vanlige gjær østrogen test, bioluminiscent gjær østrogen (BLYES) analyse15. Ved hjelp av denne linjen har de beskyttende effektene av beta-cyklodekstrin mot zearalenon-indusert toksisitet blitt bekreftet ved hjelp av kjemiske blandinger20.
I en fersk rapport ble in vivo bruk av den transgene linjen demonstrert ved hjelp av to estrogenic zearalenone (ZEA) metabolitter, α- og β-zearalenol (α-ZOL og β-ZOL)25. Protokollen baseline er hensiktsmessig å studere estrogenic effekten av flere forbindelser eller miljøprøver på Tg (vtg1:mCherry) embryoer.
